Как устроен волос, что мы с ним делаем, и как дальше жить? – 4fresh блог
Волос — это шишка. Только очень длинная. И это не шутка. Он действительно по своему устройству очень напоминает еловые шишки.
Медула
У волоса есть внутренний стержень — медула. Очень загадочная часть. Единственное, что точно можно про нее сказать, это то, что она состоит из пористого вещества, которое очень сильно напоминает губку. Такой непрерывный центральный стержень точно есть в волосах всех животных. А вот у человека медула может быть непрерывной, а может быть прерывистой. А может ее вообще не быть. Есть предположение, что именно медула отвечает за функцию «шубы» на голове — защищает от перегрева и переохлаждения. Пузырьки воздуха внутри нее создают тепловую прослойку, которая позволяет поддерживать правильный температурный режим.
С косметикой медула не взаимодействует никак, потому что ее наличие/отсутствие и целостность/прерывистость абсолютно никаким образом не сказываются на внешнем виде и красоте волоса.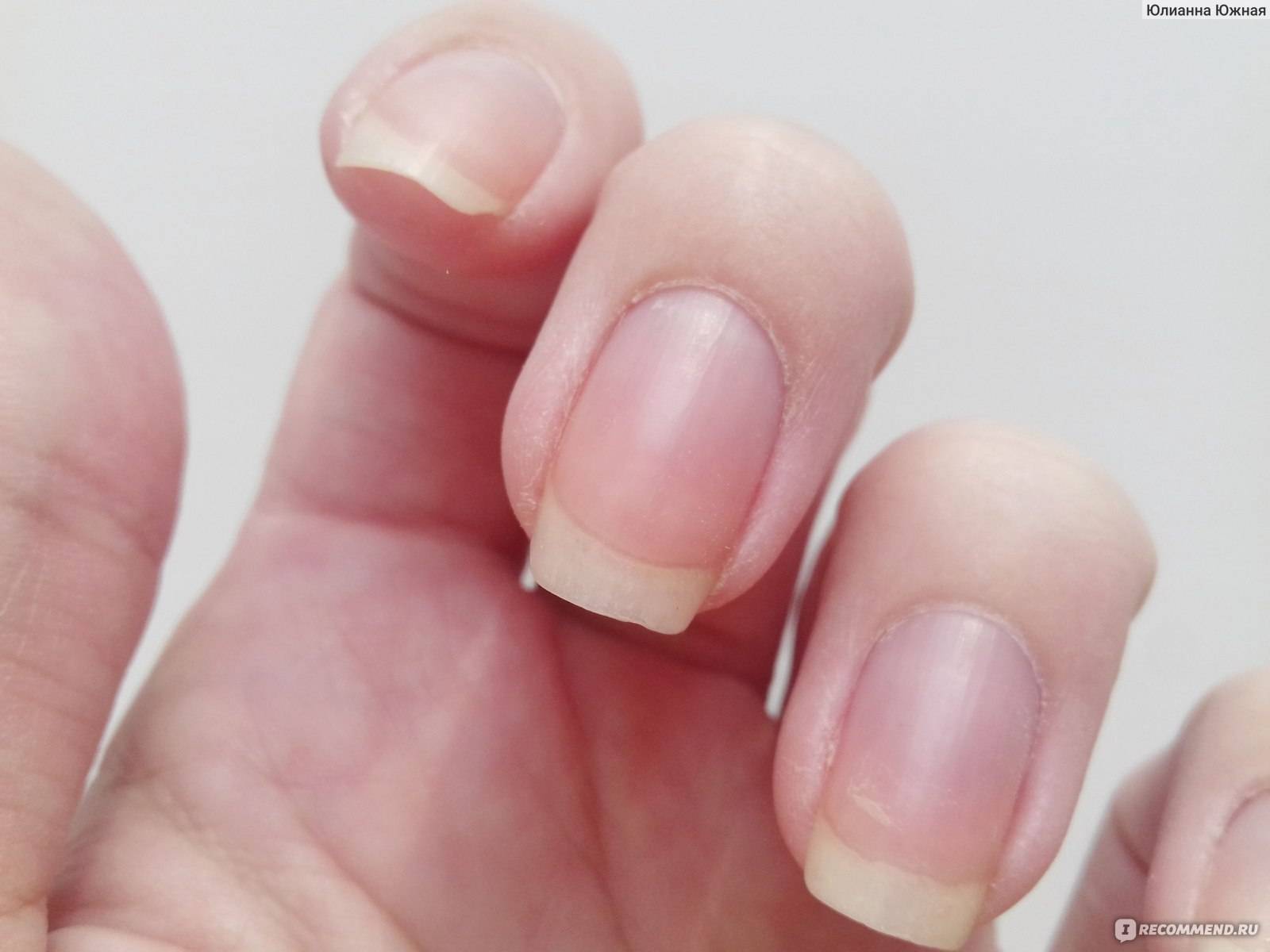
Кортекс
Второй слой волоса — это наше все — кортекс. Это основное внутреннее наполнение волоса и именно он отвечает за эластичность, прочность и цвет нашей гривы.
1) Эластичность и прочность. Кортекс по своему строению напоминает очень сложный канат — он состоит из большого количества кератиновых волокон, переплетенных между собой. Если мы разрежем такой канат, то увидим, что он состоит из более тонких канатов, а каждый тонкий канат состоит из веревочек, веревочки в свою очередь из ниточек, ниточки из волокон.
Самое тонкое волокно в волосе — это кератин — длинная белковая молекула, которая имеет форму спирали. Два кератиновых волокна свиваются друг с другом в одну суперспираль и дальше упаковываются в сложную матрешку из оболочек.
Чтобы кератиновые волокна как можно крепче держались друг за друга, они скреплены специальными мостиками, которые придают прочность волосу и… отвечают за то, будут ли виться наши волосы.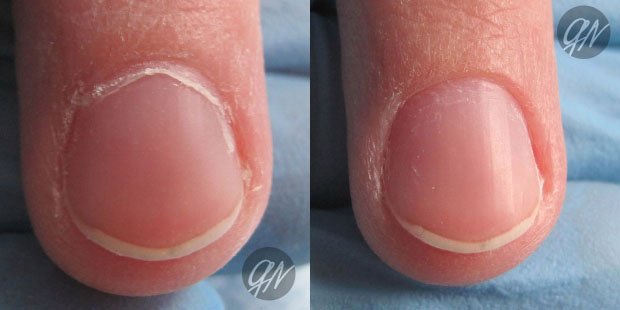 Чем сильнее обвиты волокна кератина друг об друга, и чем больше этих перемычек, тем кудрявее волосы.
Чем сильнее обвиты волокна кератина друг об друга, и чем больше этих перемычек, тем кудрявее волосы.
2) Цвет. Цвет волос определяется количеством в кортексе меланина и пузырьков воздуха.
Сам меланин бывает нескольких видов и цветов:
- Эумеланин. Это большие темные гранулы, которые цепляются за волокна кератина. Чем больше этого эумеланина, тем темнее волос. У блондинов и рыжих его нет. Внутри гранул эумеланина находится синий, красный и желтый пигмент. Синий меланин дает серые холодные оттенки и цвет воронового крыла. К сожалению, этот пигмент быстрее всего распадается на солнце, и волосы со временем выгорают и становятся каштановыми с рыжиной, т.к. в них остается меланин только красного и желтого цветов. Если мы уподобимся древним римлянкам или венецианкам средних веков и продолжим сидеть на солнце, раскинув волосы по специальной шляпе, то со временем они поменяют цвет еще сильнее и станут бледно-золотисто-рыжими.

Золотисто-рыжий цвет волос называется «венецианским». В средние века тоже была мода на блондинок, но не было перекиси водорода. Поэтому, это был максимально светлый оттенок, до которого могли выгореть темные итальянские волосы. И если внимательно присмотреться к знаменитой Венере, выходящей из пены, то можно предположить, что Ботичелли не смог добыть для себя натурщицу скандинавских кровей, а рисовал итальянку.
- Феомеланин. Это маленькие молекулы меланина красновато-желтого цвета, которые хаотично рассыпаны по кортексу. У ярких рыжих волос этого меланина очень много, а у светло-рыжих и блондинистых — мало. И чем его меньше, тем более светло-золотой оттенок приобретает волос.
Пузырьки воздуха тоже есть внутри кортекса. Зачем? Вопрос еще открыт, но чем их больше, тем сильнее они разбавляют цвет, и он становится светлее по тону. В седых волосах вообще нет никакого меланина, но есть эти самые воздушные пузырьки.
А еще пузырьки волоса отвечают за пористость волос, или за способность волос набирать в себя воду и прочие субстанции, которые попадают на волосы. Обладательницы такого типа волос обычно не пользуются маслами, т.к. эти волосы их слишком сильно впитывают и выглядят не аккуратно.


Кутикула
Ну и наконец, переходим к шишке. А точнее к верхнему слою волоса — кутикуле. Она состоит из 9-10 слоев кератиновых чешуек, которые укладываются одна на одну и составляют броню над хрупким кортексом. Чем плотнее подогнаны чешуйки кутикулы, тем сложнее сломать волос, тем он блестящее и эластичнее. Все потому, что плотный панцирь лучше отражает свет и жестче на излом.
Чем старше становится волос, тем меньше слоев чешуек на нем остается. С каждым прожитым годом их облетает все больше и больше, и как только облетают последние, то обнажается структура кортекса. Секущийся кончик волоса означает, что мы любуемся на наши кератиновые волокна, и что волос в этом месте может спокойно сломаться (вспоминаем про хрупкость кератина).
Кератиновые чешуйки в кутикуле прозрачные и не дают цвета. Но их можно окрасить разными поверхностными оттеночными красителями. Почему оттеночными? Потому что прозрачная чешуйка приобретает цвет, но сохраняет полупрозрачность и через нее светится основной пигмент волоса.
Но это еще не все. Кутикула отвечает за увлажненность волоса. Дело в том, что чешуйки не просто так прилегают одна к одной. Они склеены смесью липидов, очень похожих на липиды рогового слоя в коже. И у них точно такая же функция — препятствовать испарению воды из внутренних слоев. Есть липидная прослойка — внутренняя влага волоса запечатывается, и он сохраняет свою воду, нет — волос высыхает. Вода находится в кортексе как в свободном виде — между волокнами кератина, так и в связанном виде — цепляется за волокна. Если воды нет, то меняются физические и химические свойства кератина, и волос становится более ломким и слабым, а волосы на ощупь — жесткие и соломенные.
Чем старше становится волос, тем меньше липидов остается под чешуйками кутикулы. Чем дальше, тем больше чешуек отваливаются под действием внешних факторов, тем тоньше защитный слой кутикулы, тем сильнее сохнут волосы.
Чем дальше, тем больше чешуек отваливаются под действием внешних факторов, тем тоньше защитный слой кутикулы, тем сильнее сохнут волосы.
Именно поэтому обладательницы длинных волос так озабочены состоянием их кончиков. Они действительно будут сухими, если не предпринимать решительных мер.
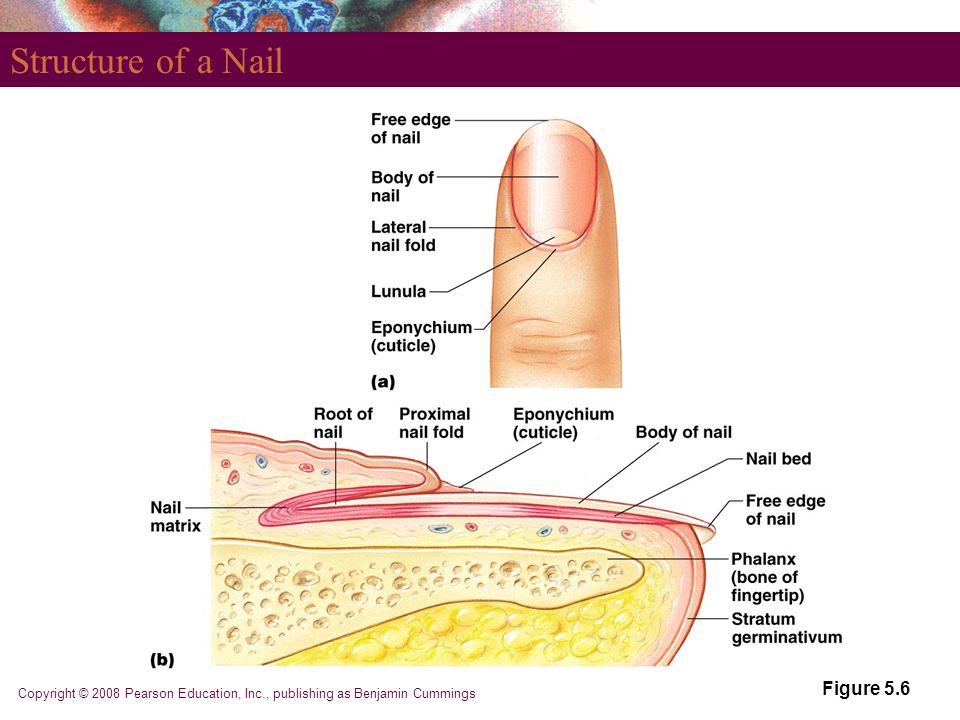
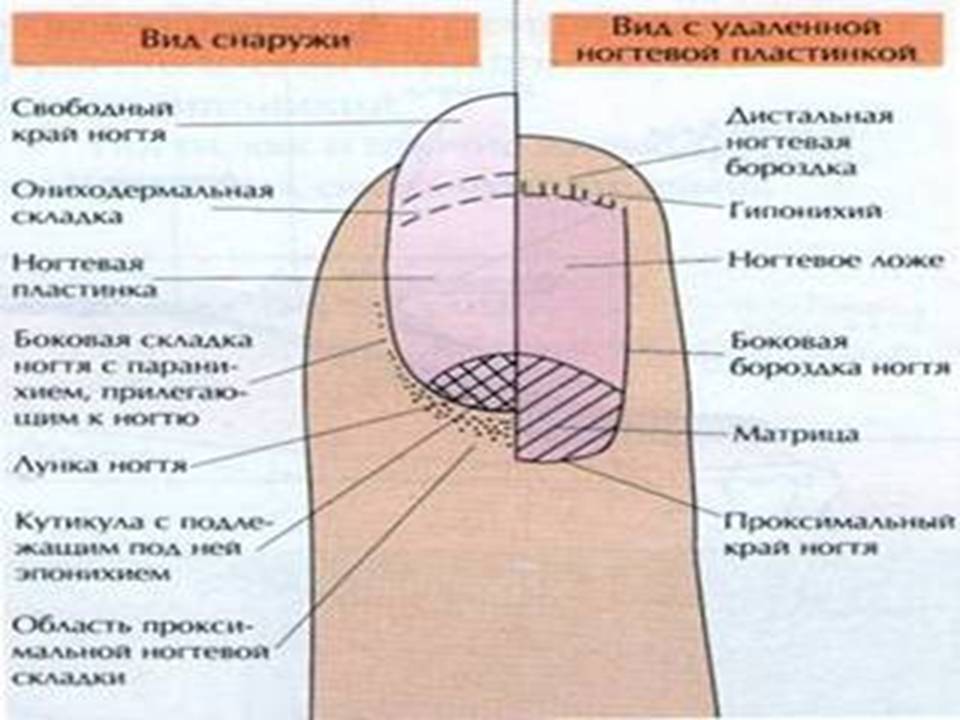
удаление и инструменты, способы ухода и лечения при воспалении
Содержание:Проксимальный валик или кутикула — это кожица, которая располагается в нижней части ногтевой пластины и состоит из плотного эпителиального валика, являющимся естественным продолжением матрикса защищающего зону роста ногтя от воздействия различных химических соединений и болезнетворных микроорганизмов (эпонихия), разрастающегося в тонкую, сросшуюся с ногтем кожистую пленку, состоящую из омертвевших клеток (птеригий).
При пересыхании и неправильном уходе кожица растрескивается, образуются отслоения и надрывы эпидермиса — заусенцы, что причиняет боль и ощущения дискомфорта. Кутикула требует особого внимания, чтобы предотвратить проблемы, следует тщательно ухаживать за ногтями. Много споров ведется о том, нужно удалять кутикулу или нет.
Кутикула требует особого внимания, чтобы предотвратить проблемы, следует тщательно ухаживать за ногтями. Много споров ведется о том, нужно удалять кутикулу или нет.
В любом случае следует понимать, что, делая маникюр, нельзя не только срезать, но и травмировать эпонихий, являющийся живой тканью и выполняющий следующие функции:
- Барьерную. Кожистый околоногтевой валик препятствует попаданию в корневую зону ногтя (матрикс) повреждающих факторов (воздуха, воды, бытовой химии, бактерий, грибов), тем самым предотвращает возникновение инфицирования и воспаления, а также деформации ногтя.
- Питательную. Через капилляры, располагающиеся в эпонихии, ноготь обеспечивается всеми необходимыми питательными веществами, нутриентами и микроэлементами, а также обогащается кислородом.
Удалять или сдвигать необходимо птеригий, ороговевший слой, который при недостаточном уходе не только может стать основной причиной образования заусенцев и повреждения околоногтевых живых тканей, но при сильном разрастании затрудняет питание ногтевой пластины и придает руке неопрятный вид.
Выбор способа ухода во многом зависит от вида кутикулы:
- Эпонихий плавно переходит в птеригий, полностью отсутствует разделяющая борозда или линия. Край кожицы плотный, упругий, эластичный, сильно сросшийся с ногтевой пластиной, трудно приподнимается и сдвигается. Обрезной маникюр в этом случае не желателен, так как из-за отсутствия видимой разделительной линии, часто повреждается живая ткань в виде порезов и трещин. Чтобы аккуратно вырезать такую кутикулу, нужно использовать маникюрные ножнички с закругленными концами и оставлять немного сдвинутого эпителия не обрезанным.
- Тонкая пленка на ногте, значительно ниже околоногтевого валика, с четкой разделительной бороздой. Хорошо поддается размягчению кератолитическими средствами и удалению при помощи кюретки или триммера.
- Жесткая, огрубевшая, выступающая над эпителиальным валиком кутикула, часто с трещинами и заусенцами. Для ее удаления используют кусачки — накожницы. На форуме специалистов маникюра есть информация, что кожицу такого типа можно в промежутках межу обрезанием смазывать бальзамом флекситолом, который способствует ее размягчению.
 На форуме специалистов маникюра есть информация, что кожицу такого типа можно в промежутках межу обрезанием смазывать бальзамом флекситолом, который способствует ее размягчению.
На форуме специалистов маникюра есть информация, что кожицу такого типа можно в промежутках межу обрезанием смазывать бальзамом флекситолом, который способствует ее размягчению.Удаление кутикулы: основные инструменты и лучшее средство для удаления
Удаление кутикулы является одним из важнейших этапов маникюра, без которого невозможно привести ручки в аккуратное и ухоженное состояние.
Кроме того, данная процедура стимулирует нормальный рост ногтей, предупреждает врастание кожицы в ногтевое ложе, а также ее надрывы, растрескивание, образование заусениц на пальцах рук.
Существует два типа удаления кутикулы: вырезание или сдвигание. Первый вариант является отличительной особенностью обрезного или классического маникюра, второй — европейского. Оба способа ухода за ногтями можно выполнять в домашних условиях или в салоне, но все-таки обрезной вариант лучше доверить специалисту.
Обрезной маникюр включает в себя:
- Подготовительный этап перед удалением околоногтевого омертвевшего слоя эпителия. Кисти рук следует погрузить на 15 — 20 минут в теплую воду, в которую можно добавить несколько капель эфирного или немного оливкового масла. Грейпфрутовое масло для ногтей поможет укрепить ногти и придать им утраченный блеск, лимонное отбелит ногтевую пластину, жожоба увлажнит кожу рук, а чайное дерево снимет воспаление и окажет дезинфицирующее и защитное действие.
- Затем следует аккуратно высушить руки мягкой салфеткой или полотенцем. Альтернативой ванночки является нанесение специального средства для размягчения кутикулы — кератолитика.
- Аккуратно при помощи пушера кожицу отделяют от ногтевой поверхности и максимально ее приподнимают. Эпидермис должен легко отделяться, не стоит сильно надавливать или царапать.
- Так называемый удалитель кутикулы выбирают в зависимости от вида кутикулы и удобства применения. Это могут быть маникюрные ножницы, триммер или специальные кусачки. Саму манипуляцию обрезания следует проводить не спеша, плавными режущими движениями, стараясь не зацепить живые ткани ногтевого валика. В идеале не должно быть порезов и крови. После процедуры нужно обязательно нанести на место среза смягчающее масло для кутикулы, которое не только предотвратит пересыхание, но и будет выполнять защитную функцию.
- После процедуры инструмент для кутикулы необходимо обязательно стерилизовать, обрабатывать антисептиком или ультрафиолетовыми лучами. Это предупредит не только нагноение пальца, но и проникновение в организм таких опасных инфекций, как ВИЧ и гепатит С.
 Кисти рук следует погрузить на 15 — 20 минут в теплую воду, в которую можно добавить несколько капель эфирного или немного оливкового масла. Грейпфрутовое масло для ногтей поможет укрепить ногти и придать им утраченный блеск, лимонное отбелит ногтевую пластину, жожоба увлажнит кожу рук, а чайное дерево снимет воспаление и окажет дезинфицирующее и защитное действие.
Кисти рук следует погрузить на 15 — 20 минут в теплую воду, в которую можно добавить несколько капель эфирного или немного оливкового масла. Грейпфрутовое масло для ногтей поможет укрепить ногти и придать им утраченный блеск, лимонное отбелит ногтевую пластину, жожоба увлажнит кожу рук, а чайное дерево снимет воспаление и окажет дезинфицирующее и защитное действие. В идеале не должно быть порезов и крови. После процедуры нужно обязательно нанести на место среза смягчающее масло для кутикулы, которое не только предотвратит пересыхание, но и будет выполнять защитную функцию.
В идеале не должно быть порезов и крови. После процедуры нужно обязательно нанести на место среза смягчающее масло для кутикулы, которое не только предотвратит пересыхание, но и будет выполнять защитную функцию.Особенностью европейского маникюра является использование специального химического препарата. Лучшее средство для удаления кутикулы, наиболее безопасное и эффективное для домашнего применения считается гель на основе фруктовых кислот, выпускающийся во флаконе с кисточкой.
- Средство необходимо нанести на кожицу на 3 — 5 минут (одновременно обрабатывают не более трех ноготков).
- Затем апельсиновой палочкой аккуратно сдвигают ороговевшие размягченные ткани вместе с остатками препарата.
- Процедуру повторяют для всех пальцев. После чего обязательно тщательно моют руки с мылом.

Чтобы кутикула росла медленнее, рекомендуется ежедневно втирать в кожицу у основания ногтя специальное средство «Авоплекс» или любое растительное масло. После размягчения эпителиальной ткани, ее следует сдвинуть при помощи апельсиновой палочки или пушера.
Регулярное использование данного метода позволяют сохранить красивый, ухоженный вид ногтя на более длительный период.
Европейский маникюр также замедляет отрастание птеригия, а если делать его каждые 2 недели и в течение периода между процедурами ежедневно смягчать и отодвигать кожицу, то можно обойтись без обрезного маникюра совсем.
Особого ухода требует очень сухая кутикула. Предупредить и устранить проблему помогут следующие советы:
- Область вокруг ногтя нуждается в регулярном пилинге. Отшелушивание омертвевшего эпидермиса следует производить при помощи готового косметического скраба, разработанного специально для этой зоны. Можно также приготовить средство для бережного пилинга в домашних условиях. Для этого нужно взять в равных пропорциях оливковое масло, воду и сахар. Тщательно перемешать, и нанести на кожу вокруг ногтевой пластины. После этого средство осторожно смывают тёплой водой, пальцы высушивают салфеткой.
- Нанесение крема на руки и область вокруг ногтей, в том числе должно быть обязательной ежедневной процедурой. Рекомендуется делать это не реже двух раз в день, а если руки часто находятся в воде или сталкиваются с повреждающими факторами, то намного чаще. Крем нужно выбирать специально разработанный для сухой кожи. Главным условием является наличие в его составе натуральных эфирных масел герани, грейпфрута, лаванды, глицерин и витаминов А и Е.
- Периодически следует использовать увлажняющие маски и ванночки (не реже раза в неделю), а при очень сухой коже ежедневно. Для этой цели подойдут растительные масла, мед, измельченный лист алоэ. Волшебными свойствами обладает состав, приготовленный из 1 чайной ложки абрикосового масла и 4 чайных ложек ланолином, смешанных до однородной массы на водяной бане; с добавлением 2 −3 капель эфирного масла.
- Соблюдая все перечисленные правила, можно стать обладательницей шикарного маникюра, фото которого станет украшением любого модного журнала.
 Можно также приготовить средство для бережного пилинга в домашних условиях. Для этого нужно взять в равных пропорциях оливковое масло, воду и сахар. Тщательно перемешать, и нанести на кожу вокруг ногтевой пластины. После этого средство осторожно смывают тёплой водой, пальцы высушивают салфеткой.
Можно также приготовить средство для бережного пилинга в домашних условиях. Для этого нужно взять в равных пропорциях оливковое масло, воду и сахар. Тщательно перемешать, и нанести на кожу вокруг ногтевой пластины. После этого средство осторожно смывают тёплой водой, пальцы высушивают салфеткой. Волшебными свойствами обладает состав, приготовленный из 1 чайной ложки абрикосового масла и 4 чайных ложек ланолином, смешанных до однородной массы на водяной бане; с добавлением 2 −3 капель эфирного масла.
Волшебными свойствами обладает состав, приготовленный из 1 чайной ложки абрикосового масла и 4 чайных ложек ланолином, смешанных до однородной массы на водяной бане; с добавлением 2 −3 капель эфирного масла.Уход за кутикулой в домашних условиях: как ухаживать правильно
Уход за кутикулой в домашних условиях удобно осуществлять при помощи специальной палочки, изготовленной из апельсиновой древесины. Именно таким инструментом пользуются приверженцы европейского маникюра.
Данный материал имеет оптимальное сочетание плотности и мягкости, что предотвращает травмы нежной кожицы вокруг ногтя. Прочная структура препятствует расслоению прибора и исключает вероятность появления занозы. Кроме того древесина обладает выраженными антисептическими свойствами.
Палочка для удаления кутикулы с одной стороны имеет заостренный край, а с другой плоский. Вернуть необходимую форму инструменту можно при помощи обычной абразивной пилочки.
Вернуть необходимую форму инструменту можно при помощи обычной абразивной пилочки.
В салоне красоты палочку можно использовать только разово, после чего она выбрасывается, так как не подлежит дезинфекции.
Основное предназначение инструмента — удаление кутикулы. После предварительного размягчения ороговевшего эпителиального слоя на поверхности ногтя, при помощи плоского конца апельсиновой палочки следует аккуратно поддеть кожицу, отделить ее от ногтевой пластины, а затем тщательно сдвинуть. После этого омертвевшие клетки эпителия можно удалить острым краем.
Кроме этого палочку используют для:
- очищения свободного края ногтя от загрязнения;
- нанесения стразов, элементов аппликации, блесток, а также для росписи лаком;
- снятия размягченного акрила, наклеивания типсов, формовки ногтя при гелевом и акриловом наращивании.
Для того чтобы ногти выглядели привлекательно, необходимо правильно ответить на вопрос, как ухаживать за кутикулой. Метод должен соответствовать ее типу и состоянию.
Метод должен соответствовать ее типу и состоянию.
Тонкую нежную кожицу внизу ногтя следует обрабатывать очень осторожно, так как существует повышенный риск травмировать ее.
Для ее удаления лучше использовать биологический метод, который заключается в размягчении эпителиальной ткани при помощи специального химического препарата. При помощи апельсиновой палочки нужно отделить кутикулу от ногтя, затем нанести гель для удаления на 0,5 — 1 минуту (если в инструкции нет других указаний). Удалить ороговевшие клетки эпителия можно при помощи ватной палочки.
Более плотную кутикулу, после ее размягчения, можно попробовать плоским краем апельсиновой палочки, пушера или триммера. Для смягчения такого типа кожицы следует ежедневно наносить на нее утром и вечером специальное средство по уходу за кутикулой или любое растительное масло. Это не только повысит эластичность эпителия, но и замедлит его нарастание на ноготь.
Труднее всего справиться с толстой ороговевшей кутикулой. Для ее удаления обычно используют щипцы или кусачки, а кератолитик применяют для предварительного размягчения кожицы. Иногда для того, чтобы привести околоногтевой валик в нужное состояние, требуется соляно-содовая или масляная ванночка, в которую пальчики опускают на 20-30 минут.
Уход за кутикулой не должен заключаться только в ее удалении или сдвигании, но и в ежедневном бережном отношении: регулярном нанесении на околоногтевую область крема, правильном питании, защите рук от агрессивных веществ с помощью перчаток. Только так можно предотвратить образование заусенец, появление микротрещин и воспалений.
Воспаление кутикулы и методы его лечения
О том, что околоногтевой валик воспалился, свидетельствуют следующие симптомы: покраснение, боль, отечность, повышенная чувствительность. В более тяжелых случаях паронихий или панариций, которые сопровождаются гнойными выделениями, подергиванием, покалыванием.
Чаще всего основной причиной патологического процесса является:
- заусенец, который не был удален правильным образом;
- слишком тонкая и легкоранимая или пересушенная и огрубевшая, склонная к растрескиванию кутикула;
- использование не обработанного инструментария для проведения маникюра;
- некоторые заболевания, в том числе грибковые поражения ногтя, аллергические дерматиты;
- различные травмы (ушиб, порез и т. д.).
Воспаление кутикулы лечение зависит от его причины и выраженности симптомов заболевания.
Если ткани вокруг ногтя просто покраснели, гной отсутствует, то достаточно будет обработать пораженную область антисептическим раствором (Йоддицерином или Фукорцином), предварительно аккуратно, под корень, срезав заусенцы, при их наличии. При необходимости место повреждения можно заклеить бактерицидным пластырем. При наличии какого-либо заболевания, вызвавшего проблему, в первую очередь следует вылечить первопричину воспаления. Лучше не заниматься самолечением и обратиться к специалисту.
Избавиться от микоза или аллергии не так просто, как обещает реклама. Врач обычно одновременно курсом назначает противогрибковые или противоаллергические препараты для внутреннего и наружного применения. Положительный результат терапии возможен только при четком соблюдении всех рекомендаций.
Если воспаление приняло более серьезный оборот, появились гнойные выделение лечение кутикулы в домашних условиях практически не возможно. В большинстве случаев, требуется хирургическое вмешательство, которое направлено на вскрытие нарыва.
После этого рану хорошо промывают антисептиками (Перекисью водорода, Хлоргексидином биглюконатом или Мирамистином). Если рана глубокая, для оттока выделений ее дренируют. Затем накладывают повязку с бактерицидной мазью, лучше, если кроме антибиотика в ее состав входит вещество стимулирующее регенерацию тканей (Левомеколь, Левосин). Перевязки нужно делать под контролем врача 1 — 2 раза в день, до полного заживления. При поражении глубоких слоев тканей или при отсутствии эффекта от местной терапии назначают антибиотики внутрь или в виде инъекций (Левомицетин, Цефалексин).
Если причиной постоянного воспаления кутикулы является ее состояние, то следует обратить внимание на мероприятия, которые ее восстановят. Нужно:
- Сбалансировать дневной рацион, включить в его состав все необходимые для нормального формирования кожи компоненты.
- Регулярно делать маникюр, причем соблюдать все правила качественной процедуры.
- Часто (не менее 2 раз в день) наносить на руки хороший увлажняющий крем.
- Пить достаточно жидкости в течение дня.
- Всегда использовать защитные перчатки во время работы в саду и огороде, контакте с бытовой химией.
- На ночь намазывать кожицу вокруг ногтя оливковым маслом и одевать хлопчатобумажные перчатки.
- Пить витаминные комплексы, содержащие ретинол, токоферол, биотин и некоторые другие нутриенты.
Автор: Александр Шаповалов
Структура волоса – часть 1: Кутикула
18 января, 2019
Рисунок №1: Структура волос с кутикулой, кортикальными клетками, макрофибриллами, промежуточными филаментами, протофибриллами, протофиламентами и димерами.
В наше время ежедневный уход за волосами – необходимость. Чтобы сохранить и улучшить состояние волос, нужно использовать средства для ухода за ними – кондиционеры для волос, шампуни, средства для укладки волос, продукты для поддержания формы волос и так далее. При выборе надо учитывать два фактора. Первое – для правильного выбора средства для ухода за волосами всем людям, начиная от обычных потребителей, заканчивая парикмахерами и стилистами, работающими в салонах красоты, необходимо знать как устроены волосы. Второе – практически все средства вызывают изменения в структуре волос.
Начнем с того, что у волос очень сложная структура, но если основываться на базовых понятиях, которые были собраны простым методом исследования волос, то их структура становится понятной всем людям. В целом предмет изучения структуры волоса очень интересен для ученых, и именно из-за этого в последние годы было сделано так много открытий в этом направлении. Благодаря этим исследованиям можно понять, что шутить со здоровьем волос нельзя, и нужно ответственно подходить к уходу за ними, а также к их лечению (если оно требуется), дабы избежать повреждений в будущем. Одним из главных фактов, полученных в результате исследований, который стоит запомнить это то, что у людей разных национальностей структура волос может кардинально различаться. К примеру, как у людей с Африканского континента и из Азии.
Начиная описывать структуру волос важно сказать, что они состоят из водонерастворимого белка, известного как кератин, который в свою очередь состоит из множества аминокислот. По сути волос имеет в своем составе три части: первая – кутикула, вторая – кора, а третья – медулла. Все эти три части можно было бы описать в одном блоге, но тогда бы он вышел скучным и очень длинным, поэтому я разобью их описание на три части. И так, первый блог будет посвящен кутикуле.
Кутикула
Кутикула – это защитный слой, который образует внешнюю поверхность волос. Она состоит из нескольких слоев, преимущественно от 6 до 10, склеенных вместе с клеточным мембранным комплексом (CMC). Однако на завитой части волоса кутикула имеет всего лишь два слоя.
Рисунок №2: Слои кутикулы
Диаметр волосяного волокна составляет около 80 микрон (мкм). Чтобы понять насколько эта малая единица измерения достаточно узнать, что микрон составляет одну миллионную часть метра в длину. У всех типов волос будет свой диаметр. Так, к примеру, у мелких волос будет диаметр от 50 до 60 микрон, средние волосы имеют диаметр от 61 до 80 мкм, а грубые толстые волосы – диаметр от 81 до 100 мкм. На данный факт стоит обратить особое внимание т.к. для каждого типа волос подходят свои средства по уходу за ними.
Каждый слой кутикулы имеет длину около 60 микрон (мкм) и толщину 0,5 мкм. Длина области перекрытия одного слоя кутикулы над другим слоем кутикулы составляет около 5 мкм . Слишком вьющиеся волосы имеют различное количество слоев кутикулы вдоль протяженности волоса. На частях, где волосы тонкие или плоские, вдоль малой оси волос находятся всего лишь один или два слоя, а вдоль большой оси, где толстые волосы, имеется от 6 до 10 слоев. Основная и малая ось поперечного сечения волос обобщаются подзаголовком «Эллиптичность волокна». Кутикулы также богаты аминокислотой, известной как цистеин и некоторыми жирными кислотами. Внешняя часть кутикулы волос состоит из жирной кислоты, называемой 18-МЭА (18-метиликоносановая кислота), также известной как F-слой. Все 18-МЭА находятся на самой верхней поверхности кутикулы. Структура F-слоя представлена на рисунке ниже.
Рисунок №3: Структура F – слоя
Мембранный комплекс клеток (CMC) кутикул: каждый слой кутикулы приклеивается к следующему слою кутикулы с помощью материала, называемого межклеточным цементом или клеточным мембранным комплексом (CMC) и его межклеточными пространствами.
Типы CMC: Существует три типа клеточных мембранных комплексов.
Первый тип CMC – находится только между кутикулами. Второй тип CMC находится между внутренним слоем кутикулы и кортикальными клетками. Третий CMC находится между самими кортикальными клетками. Все эти три СМС несколько отличаются по своим композициям и объясняются далее.
Существенная роль CMC между кутикулами – это сцепление одного слоя кутикулы с другим. Схематическое изображение CMC кутикулы показано на рисунке 4.
Рисунок №4: Схематическое изображение CMC кутикулы
Внешние бета-слои составляют 5 нанометров (нм) и состоят из липидов. Центральный «Дельта» слой составляет 15 нанометров (нм).
Второй тип СМС – именно тот, который обеспечивает сцепление между самым внутренним слоем кутикулы и внешними кортикальными клетками коры. Эта CMC легко повреждается при обработке растворителями, такими как вода или водосодержащими моющими средствами, а также агрессивными химикатами. В сравнении СМС «кутикула к кутикуле» получает практически такой же урон, однако не сравним с малым уроном который получает СМС типа «кортико-кортикальной».
Третий тип CMC – это клеточный мембранный комплекс, который удерживает клетки коры склеенными. СМС кортикально-кортикальной клетки более легко повреждается с помощью растворителей по сравнению с CMC 1 типа и CMC 2 типа. Все эти три СМС несколько отличаются по своему взаимодействию с растворителями .
Пространства внутри CMC являются предпочтительным путем проникновения и диффузии многих веществ в кору волос. Структура этих пространств в CMC пока неизвестна. Реактивные химические вещества, такие как щелочи (гидроксиды натрия и гидроксиды гуанидина), восстановители (тиогликолят аммония) и окисляющие агенты (перекись водорода), нападают на липиды CMC. Деградация CMC между кутикулами может привести к разрыву слоев кутикулы, быстро ослабить ее и она больше не сможет выполнять эффективную роль в качестве защитного средства для коры волос.
Структура кутикулы
Структура самой кутикулы довольно сложна и далее подразделяется на четыре части, такие как Эпикутикула, А-слой, Экзокутикула и Эндокутикула, как показано на рисунке 5.
Рисунок №5: Части кутикулы: Эпикутикула, А-слой, Экзокутикула и Эндокутикула
Эпикутикула – очень тонкая мембрана, которая покрывает кутикулу и является внешним слоем, как показано на рисунке 4. Она составляет от 5 до 7 нанометров (нм) толщиной. Слой очень гидрофобен по своей природе, это означает что слой не переносит воду. F-слой (18 – MEA) не очень устойчив к щелочам, окислителям и протеолитическим агентам и при длительном воздействии, существенные химические изменения имеют место и в эпикутикулярном слое, тем самым делая волосяные волокна более пористыми, увеличивая трение между фибриллами волос. Волокна африканского происхождения имеют менее 18-МЭА на поверхности волосяных волокон по сравнению с кавказскими необработанными волосами. Вот и подтверждение примеру, о различии структуры волос у людей разных национальностей приведенного мною в начале блога.
A-слой: слой «A» существует ниже слоя эпикутикулы. Слой «A» и слой экзоцитиля образуют около двух третей масштабной структуры кутикулы. Они очень богаты содержанием цистеина. В слое «А» содержится около 35% цистеина. Слой «А» противостоит физическим воздействиям, таким как трение во время расчесывания, мытья и сушки; и химическим воздействиям, таким как щелочи, восстановители, окислители и протеолитические ферменты, которые в противном случае могли бы разрушить целостность волосяного волокна без кутикулы.
Экзокутикула: подслой под слоем «А» называется экзокутикулы и содержит около 15% содержимого цистина, как показано на рисунке 4. Экзокутикулы не имеют фибриллярной структуры. Слой «А» на самом деле является частью экзокутикулы, которая делятся на слой «А» и «В», именно слой «В» обычно называют экзокутикула.
Слой эндокутикулы находится рядом с слоем экзокутикулы и содержит низкое содержание цистеина – около 3%. Это механически самый слабый компонент структуры кутикулы. Существует большой пробел в содержании цистеина между эндокутикулой и «A – Слоем». Из-за очень низкого присутствия содержимого цистеина в слое эндотелия, он имеет очень мягкую и поддающуюся деформации структуру, а также значительно больше разбухает в воде, чем слои экзокутикулы. Явное выступание кутикулы во влажном состоянии связано с тем, что слой эндокутикулы набухает при смачивании водой. Переплетение волокон волос гораздо больше во влажном состоянии, и это связано с необычайным набуханием эндокутикулы. Слой эндокутикулы может легко повреждаться протеолитическими агентами и другими реакционноспособными химическими веществами, к примеру щелочами. Особым преимуществом слоя эндокутикулы может быть то, что он может обеспечить некоторую защиту, создавая подушку под более жестким внешним слоем экзокутикулы из сил, воздействующих на поверхность волос. Поэтому предполагается, что роль кутикулы заключается в том, чтобы противостоять проникновению реакционно способных и не реакционно способных химических веществ в кору волос. Эта защитная роль кутикулы является благословением для долговременной жизни волокон волос. Кутикулы волосяных волокон также противостоят изгибу волосяного волокна на 74%, из которых 66% этого сопротивления приходится на экзокутикулу, а 8% – на эндокутикулу.
Кутикула
Определение «Кутикула» в Большой Советской Энциклопедии
Кутикула (от лат. cuticula — кожица), 1) у животных — плотное образование на поверхности клеток эпителиальной ткани. У беспозвоночных — производное клеток однослойного покровного эпителия (см. Гиподерма). Выполняет главным образом защитную и опорную функции. У червей, членистоногих и др. образует прочный покров тела. В состав Кутикула входят хитин, который вместе с минеральными веществами и белками придаёт Кутикула механическую прочность, и липиды, способствующие её водонепроницаемости. У позвоночных, которые имеют многослойный покровный эпителий, выполняющий защитную функцию, Кутикула находится преимущественно на поверхности эпителиальных клеток, выстилающих внутренние органы (кишечник, воздухоносные пути органов дыхания, части мочевых канальцев почки и мочеотводящих путей). Каждая клетка имеет свою Кутикула, представленную совокупностью микроворсинок (на поверхности одной клетки тонкой кишки человека их помещается до 2 тыс.). В Кутикула обнаружена активность некоторых ферментов (например, щелочной фосфатазы, инвертаз, мальтаз), что свидетельствует об активном участии Кутикула в функциональной деятельности органов.
2) У растений — тонкая плёнка, покрывающая эпидермис листьев и стеблей и выполняющая защитную функцию; состоит из кутина. Кутикула — бесструктурное образование, лишённое корпускулярных и фибриллярных элементов; устойчива к химическим воздействиям. Она отсутствует на погруженных в воду органах водных растений, слабо развита у растений, обитающих в тени и на сырой почве, и особенно хорошо — у растений, нуждающихся в ограничении транспирации. Гладкая и блестящая Кутикула листьев тропических растений отражает часть солнечных лучей и служит защитой от чрезмерной инсоляции. У многих ксерофитов защитные свойства эпидермиса усиливаются расположенным под Кутикула т. н. кутикулярным слоем, состоящим из смеси полисахаридов, кутина и восков. У большинства ксерофитов в кутикулярном слое откладываются бледно-жёлтые пигменты, обеспечивающие непроницаемость клеточной стенки для ультрафиолетовых лучей.
М. Е. Аспиз.
Статья про «Кутикула» в Большой Советской Энциклопедии была прочитана 250 раз
Строение волоса и подбор процедур. Статьи компании «KERATIN-PROF Беларусь»
Немного информации о строении волос, обязательна к изучению начинающим и будущим мастерам. Также в статье Вы узнаете, как научится подбирать процедуры по типам и состоянию волос
Чтобы иметь возможность правильно подобрать составы для процедур, нужно понимать, как устроены волосы, от чего и как они разрушаются и как их можно восстановить.
Начнем со строения.
Мы будем говорить о той части волоса, которая считается мертвой, то есть наружном стержне.
В процентном соотношении волос имеет следующий химический состав:
78%-кератин
6%-липиды
1%-пигмент
15%-вода
Часть волоса, расположенная над поверхностью эпидермиса, называется стержнем. Невидимая часть, расположенная в толще кожи, называется корнем (луковицей).
Современные средства по уходу за волосами бренда эффективно работают на стержне волоса. Волос состоит из следующих составляющих:
1. Кутикула представляет собой внешнюю оболочку из ороговевших чешуек. Она служит защитой для внутренних слоев от внешних воздействий. Именно кутикула обеспечивает блеск и шелковистость волос.
2. Кортекс (корковый слой волоса) находится непосредственно под кутикулой. Он состоит из множества ороговевших веретенообразных клеток, которые можно сравнить с нитью. Эти переплетенные «ниточки» образуют микрофибриллы волоса, которые составляют крупные волокна-макрофибриллы. Чтобы миллионы микрофибрилл не распадались между собой на молекулярном уровне, существуют поперечные связи: дисульфидные, водородные и солевые.
Из-за особенностей в строении кортекса волосы могут быть прямыми и кудрявыми, которые в свою очередь наследуются генетически. В клетках этого слоя находятся пигменты, придающие волосам цвет. Кортекс также отвечает за прочность и эластичность. В данном слое происходит химический процесс выпрямления, разглаживания или окрашивания волос.
3. Медула (сердцевина) — это стержень волоса, который состоит из 2-4 слоев, не ороговевших клеток различной структуры с кератиноподобным веществом. Медула отвечает за терморегуляцию, объем и придает силу волосам. В наше время сердцевина у многих прерывистая или полностью отсутствует
Чешуйчатый слой предохраняет нижележащие клетки кортекса от пагубного действия условий внешней среды. Кутикула волоса защищает кортекс и медуллу как панцирь. Она образована перекрывающимися слоями прозрачных пластинок кератина, напоминающих чешую. Кутикула препятствует механическим и химическим воздействиям на волос. Неповрежденная кутикула хорошо отражает свет: волосы блестят, не ломаются. Средства с высоким РН (щелочные — мыло, например), расслабляют чешуйки и заставляют их открываться и пропускать жидкость внутрь кортекса. Средства с низким уровнем РН (кислые) захлопывают чешуйки, делают их плотнее и прочнее.
Вот здесь кроется первый важный момент, который нужно понимать: чешуйки кутикулы раскрываются и закрываются от изменения уровня РН!!! Например, РН воды в море чаще имеет щелочную среду, РН воды в кране может иметь щелочную среду, и большинство шампуней (особенно масс-маркет) — это щелочи. Значит, если ваши волосы подверглись воздействию щелочи, то вам необходимо обработать их кислым средством, чтобы они закрылись (например, бальзам). Волосы с раскрытыми чешуйками кутикулы очень хрупкие, легко разрушаются и, конечно же, не блестят.
Второй важный момент: если вас интересует красота волос, их блеск, то вам нужно позаботиться о состоянии кутикулы!
Кутикулу тоже можно механически повредить, например, грубым расчесыванием, горячими щипцами. В этом случае даже при обработке специальными средствами она не сможет обеспечить полную защиту кортекса, и волос начнет разрушаться. Поэтому очень важно бережно обращаться с волосами, мягко расчесывать их без резких движений, а также как можно реже подвергать их нагреванию и сушке. В этом случае рекомендуется использовать термозащитные спреи и лосьоны.
Теперь перейдем к кортексу, среднему слою, который составляет примерно 90% волоса человека. Именно в этом слое происходят изменения при окрашивании или химии. От состояния кортекса зависят сила, упругость и прочность волоса. Волос с хорошо развитым корковым слоем значительно прочнее волоса со слабо развитым слоем.
Корковый слой окружает сердцевину. Он представляет собой плотно прилегающие одна к другой клетки, продольно вытянутые и расположенные по длине волоса. При рассмотрении под электронным микроскопом видно, что эти клетки состоят из тончайших волокон, фибрилл, а те — из еще более тонких волокон, и так далее, пока не дойдем до молекулярного уровня. Здесь можно провести аналогию со строением каната, который сплетен из более тонких волокон. Волокна кортекса состоят в основном из белка — кератина. Белок состоит из длинных цепей аминокислот, которые в свою очередь состоят из химических элементов. Основными химическими элементами, из которых состоит человеческий волос являются: углерод, кислород, водород, азот и сера. Из этих элементов так же состоят кожа и ногти.
Белки — это длинные аминокислоты, соединенные между собой на концах прочными белковыми связями. Связываясь друг с другом, они образуют длинные, спиральные, сложные аминокислотные цепочки. Эти спирали образуют микрофибриллы, которые перекручиваясь, образуют волокна кортекса. Итак, мы разобрались, как волокна образуются в длину. Но что удерживает их вместе, как в тугом канате? Чтобы миллионы полипептидных цепочек не распадались, между ними также существуют поперечные связки или мостики. Связки эти удерживают волокна вместе на молекулярном уровне.
Эти связи имеют огромное значение для таких услуг, как термическая укладка, химическая завивка и химическое выпрямление волос. Основными поперечными связями являются: водородные, ионные или солевые и дисульфидные
Водородные связи легко разрушаются водой и температурой. И хотя по отдельности водородные связи очень слабые, их очень много в волосах, поэтому они дают примерно одну треть силы волоса. Водородные связи легко разрушаются при намокании волос. Это позволяет волосам легко растягиваться. При высыхании и охлаждении водородные связи восстанавливаются и фиксируют форму волос в том положении в котором волосы оказались. Так, например, завивка волос на обычных бигуди и высушивание их естественным образом приводит к разрушению водородных связей, благодаря чему, волосы из прямых становятся закрученными, но через некоторое время или под воздействием влажной среды, разрушенные водородные связи восстанавливаются и волосы снова приобретают прежнюю форму.
Ионные или солевые связи так же являются слабыми физическими связями, образующимися поперек волоса между соседними белковыми цепочками. Ионные связи зависят от pH на волосах, поэтому они легко разрушаются сильными щелочными и кислыми растворами. Но их в волосах так же много, поэтому оказывают значительное влияние на структуру и свойства волос. Солевые связи восстанавливаются при нормальном рН волос.
Дисульфидная связь — это сильная химическая связь, которая очень отличается от физических водородной и ионной. Она соединяет атомы серы двух соседних аминокислот, чтобы создать одну, объединяет две белковые цепочки в единое целое.
Хотя самих дисульфидных связей гораздо меньше, чем водородных и ионных, они очень сильные и не разрушаются водой. Разорвать дисульфидную связь можно с помощью препаратов для химической завивки и выпрямления. Нормальное температурное воздействие, как, например, при термоукладке, не разрушает дисульфидных связей. Однако очень сильные воздействия температуры, как при воздействии кипятка и некоторых высокотемпературных инструментов для укладки, нарушают эти связи. Яркий пример такого сильного воздействия — утюжки для выпрямления волос, температура которых достигает более 200 градусов.
Если дисульфидные связи были разорваны и не восстановлены химическим путем, то волос остается поврежденным. Со временем он становится хрупким, ломким, пористым, сильно обезвоженным и истонченным. Это приводит к посеченным концам, волосы тяжелее расчесываются и становятся на ощупь жесткими и сухими.
Итак, очередной вывод: если вас беспокоят сила и упругость ваших волос, то вам нужно задуматься о восстановлении кортекса волоса, то есть восстановлении дисульфидных связей (всегда используйте специальные средства при химическом воздействии или окрашивании! а также специальные шампуни и бальзамы, например, с гематином) и восстановлении волокон кортекса
Кстати, кератины в составе шампуней не способны проникнуть вглубь волоса, поэтому восстанавливают лишь частично, заполняя пустоты у внешней оболочки непосредственно под чешуйками кутикулы. При несильном повреждении волос, они могут дать восстанавливающий эффект, но при глубоких повреждения — не очень эффективны.
Кроме всего прочего в клетках коркового слоя обычно располагаются зерна красящего вещества — пигмента. При окрашивании этот пигмент разрушается и заменяется искусственным, но искусственный пигмент не является таким устойчивым, поэтому легко вымывается из кортекса волоса при мытье головы (после окрашивания постарайтесь не использовать сульфатные шампуни, это позволит дольше сохранить цвет), оставляя после себя пустоты с воздухом, что делает волос более хрупким.
Вообще, конечно, восстановление волос — это комплексная терапия, включающая в себя как лечение, восстановление, так и защиту волос от дальнейших повреждений. Она требует дополнительных мер, ухаживающих средств и большого желания и терпения.
С ЧЕГО НАЧАТЬ?
После детального изучения строения волос, нужно научиться понимать, как правильно подбирать процедуру по типу и состоянию волос клиента.
1. После знакомства с клиентом необходимо узнать у него «историю» его волос. Какие были окрашивания, какой оксид использовался. Окрашивали ли волосы хной? Были ли смывки/осветления. Делал ли клиент процедуры кератинового выпрямления/ботокса ранее? Как реагировали на процедуру волосы и сколько состав срок носки
2. Также уточняйте у клиента, не принимает ли он серьезные гормональные препараты, о беременности, о кормлении грудью
Далее проведите визуальный осмотр волос клиента, сделайте тест на степень повреждения волос (намочите волос и аккуратно растяните его). Если волосы растягиваются и не возвращаются в прежнее состояние-таким волосам не рекомендуется проводить «горячие» процедуры, т.е. процедуры с использованием утюжка, также можно проверить волосы на пористость.
В случае если волосы сильно повреждены, а именно рвутся-рекомендуется провести процедуру сос-реконструкции холодным способом. Можно использоваться составы Felps SOS, ампулы от Dr.Therapy, каутеризацию от Gold Spell и другие составы для реконструкции.Такие составы направлены именно на восстановление дисульфидных связей волоса. Эти процедуры можно делать с помощью инфракрасного утюжка, иногда требуется курс восстановительных процедур (необходимо смотреть за состоянием волос). Курс может составлять 2-5 процедур.
После курса реконструкции, когда волосы перестали рваться, а при растягивании возвращаются в прежнюю форму-можно выполнять более серьезные восстановительные процедуры с использованием утюжка.
БОТОКС ИЛИ КЕРАТИН?
В целом это довольно схожие процедуры, большинство современных составов для ботокса по компонентам близки к составам для кератинового выпрямления волос. Как правило, в ботоксах содержится большее количество ухаживающих, увлажняющих компонентов (масла, кодиционеры), чем в кератине. За счет этого составы для ботокса дают больший эффект увлажнения и восстановления волос.
Кератин в первую очередь направлен на выпрямление волос, содержит большее количество выпрямляющих компонентов. Современные составы для выпрямления не сильно уступают ботоксам в содержании увлажняющих компонентов. Поэтому при правильно подобранном по типу волос составу можно получить отличный результат выпрямления и восстановления/увлажнения волос.
На какие волосы рекомендуется делать именно ботокс:
Окрашенные на высоких оксида (блонд), волосы после смывки, осветления, других сложных окрашиваний-после оценки состояния волос. На волосы после химической завивки (также в зависимости от состояния волос). Также ботокс рекомендован на ПРЯМЫЕ от природы волосы (натуральные, окрашенные), так как такие волосы нет смысла перегружать «тяжелыми» составами для выпрямления волос, а также не подвергать волосы серьезной термообработке, как при выпрямлении волос. С ботоксом можно работать на более низких температурах.
Также ботокс можно делать на вьющиеся волосы, для структуризации кудрей, устранения пушистости, увлажнения. Важно при этом подобрать ботокс без эффекта выпрямления. Есть довольно сильные составы для ботокса, которые смогут на некоторое время выпрямить кудри.
Основной показатель выполнения процедуры кератинового выпрямления-вьющиеся волосы, которые клиент хочет выпрямить, при этом часто пользуется утюжком и регулярно травмирует их. Далее стоит учитывать состояние волос, убедиться, что волосы смогут выдержать необходимую для термообработки состава температуру (оптимальная температура утюжка при работе с кератином 210-230 градусов). В зависимости от состояния волос температура может быть ниже, но это уже может сказаться на эффекте выпрямления и сроке носки состава. Поэтому, если Вы видите, что волосы не выдержат высокую температуру-предварительно подготовьте их, сделайте более щадящую процедуру-ботокс или реконструкцию. После этих процедур волосы смогут без труда выдержать серьезную термообработку и можно сделать выпрямление волос через некоторое время.
Морфология и биология волос: правильный выбор ингредиентов
Волосы – это природное украшение любого человека. Иметь красивые, ухоженные, блестящие волосы не откажется никто. Средства ухода за волосами – огромный сегмент косметического рынка, и именно этот сегмент рынка особенно важен для российского производителя косметики. Чтобы создавать средства для волос, которые будут востребованы потребителями, недостаточно хорошо разбираться в химии. Полезно иметь представление о том, что же такое волосы с точки зрения биологии, как они устроены и как живут. Именно этим вопросам и посвящена настоящая статья.
Волос под микроскопом
Биологическая функция волос – это защита головы от перегревания, охлаждения, механических повреждений. Примерный химический состав здорового волоса таков: 3–15% воды, 6% липидов,1% пигмента,78–90% белка (кератина). Основными элементами в составе волос являются углерод (49,6%), кислород (23,2%), азот (16,8%), водород (6,4%), сера (4%). Кроме того, в состав волоса входят магний, мышьяк, железо, фосфор, хром, медь, цинк, марганец, золото.
Рисунок 1. Строение волоса
С точки зрения физиологии, волос – это «мертвое» образование. Волосы не снабжаются кровью, к ним не подходят нервные волокна и не присоединяются мышцы. Поэтому при стрижке мы не чувствуем боли, волосы не кровоточат, при их натяжении не растягивается ни одна мышца. Но, если волос – это мертвое образование, то почему волосы растут и почему, когда мы выщипываем волосы, нам больно? Дело в том, что каждый волос состоит из двух частей: видимой части (стержня волоса) и скрытого в коже фолликула. Живые клетки, которые размножаются с огромной скоростью, находятся в волосяном фолликуле, залегающем глубоко в дерме.
Волосяной фолликул – это корень волоса с окружающими его тканями и волосяно-железистый комплекс (сальная и потовая железы; мышца, поднимающая волос; кровеносные сосуды и нервные окончания) (рис.1). Мы рождаемся на свет с определенным количеством таких фолликулов, величина эта генетически запрограммирована, и изменить здесь что-либо на данный момент времени невозможно. Количество волосяных фолликулов разное у людей с разным цветом волос. В среднем, общее количество волос на голове у блондинов – 140 тыс., у шатенов – 109 тыс., у брюнетов – 102 тыс., а у рыжих – 88 тыс.. Скорость деления клеток волосяного фолликула занимает в организме человека второе место после скорости деления клеток в костном мозге. Благодаря этому волосы вырастают примерно на 1–2 сантиметра за месяц.
Каждый волосяной фолликул имеет собственную иннервацию и мускулатуру. Благодаря нервным окончаниям, волосяной фолликул обладает тактильной чувствительностью. Мышцы, примыкающие к фолликулу, сокращаясь от страха или под влиянием холода, приподнимают волосы и сжимают кожу, в результате на коже образуются «мурашки». В устье волосяного фолликула открывается проток сальной железы. Эта железа вырабатывает кожное сало, которое, выделяясь в устье волосяного фолликула, смазывает растущий волос и поверхность кожи волосистой части головы. Кожное сало создает на волосах пленку, придавая им эластичность, гладкость и водоотталкивающую способность. Активность сальных желез контролируется половыми гормонами – андрогенами. В устье фолликула поселяются некоторые бактерии – представители микрофлоры кожи. Изменения микрофлоры ведут к нарушению кератинизации и появлению перхоти.
Волосяной фолликул – «фабрика» по производству волос
Итак, волос «зарождается» в фолликуле (рис.2). Рост нового волоса начинается от волосяного (дермального) сосочка – соединительнотканного образования, находящегося в основании фолликула (волосяной луковицы). Волосяной сосочек содержит в себе кровеносные сосуды, снабжающие клетки луковицы кислородом и питательными веществами, необходимыми для размножения клеток и роста волос. Если гибнет сосочек – погибает и волос. Клетки фолликула делятся и размножаются в зоне, прилегающий непосредственно к волосяному сосочку. По мере продвижения к поверхности кожи головы фолликулярные кератиноциты постепенно теряют свои ядра, уплощаются и ороговевают, заполняясь твердым кератином. Среди клеток волосяной луковицы представлены и меланоциты, которые обуславливают интенсивность цвета волос. Чем больше меланоцитов – тем темнее волос. С возрастом активность меланоцитов падает, в результате чего волосы седеют. Оттенок волос определяется генетическими факторами и зависит от соотношения содержания двух основных пигментов: эумеланина и феомеланина (рыжие волосы). Таким образом, цвет волос зависит от комбинации двух факторов: соотношения пигментов и количества синтезирующих пигмент клеток.
Рисунок 2. Строение корня волоса
Каждый фолликул является независимым образованием со своим собственным жизненным циклом (рис.3). Жизненный цикл волоса состоит из трех стадий, его продолжительность колеблется от 2 до 5 лет. Каждый волосяной фолликул генетически запрограммирован на производство примерно 25–27 волос.
Рисунок 3. Жизненный цикл волос
Первая фаза жизненного цикла волос –анаген, когда фолликул производит тонкое волосяное волокно, которое постепенно растет и становится толстым и пигментированным. Анаген продолжается от двух до пяти лет. Следующая фаза – катаген – фаза покоя. Деление клеток фолликула замедляется и прекращается, волосяной фолликул «впадает в спячку». Волосяная луковица постепенно отсоединяется от волосяного сосочка. Эта фаза длится очень недолго – от одной недели до трех недель. И последняя фаза, завершающая жизненный путь волоса – телоген. В период телогена новый волос начинает расти, а старый выпадает. Обновление клеток прекращается приблизительно на 3 месяца (время, за которое восстанавливается связь между вновь синтезированной волосяной луковицей и волосяным сосочком, и новый волос входит в фазу анагена). Полностью отделившаяся от дермального сосочка телогеновая луковица приобретает вытянутую форму и начинает двигаться к поверхности кожного покрова волосистой части головы.
Каждый волос живет по своему «индивидуальному графику», потому разные волосы в одно и то же время находятся на разных стадиях своего жизненного цикла: 85% волос находятся в фазе анагена, 1% в фазе катагена и 14% – в стадии телогена. Когда соотношение между волосами, находящимися в фазах анагена и телогена, нарушается и в стадии выпадения оказываются более 14–15% волос, можно говорить о развитии облысения или алопеции. В норме в день мы теряем в среднем 70–80 волос в день.
Стержень волоса
Стержень волоса имеет сложную конструкцию. В центре его находится мозговое вещество, окруженное кортексом, то есть корковым слоем (около 80–90% всего объема волоса). Снаружи волоса находится кутикула, которая покрывает кортекс, будто черепицей.
Медуллярный слой (центральное мозговое вещество) – это центральная часть волосяного стержня, которая представлена у человека не во всех видах волос. Например, в пушковых волосах медулла отсутствует. Мозговое вещество заполняют пузырьки воздуха – благодаря этому волос обладает определенной теплопроводностью. Медулла не играет никакой роли в изменении как химических, так и физических свойств волоса.
Корковый слой, или кортекс – это основное вещество волоса, которое составляет от 80 до 85% его объема. Кортекс состоит из рогового вещества кератина, образованного склеенными ороговевшими веретеновидными клетками. Как и любой другой белок, кератин состоит из аминокислот, которые формируют полипептидные цепи. Эти цепи переплетаются между собой, образуя нити. Эти нити, в свою очередь, навиваясь друг на друга, создают суперспирализованную структуру: объединяясь по несколько штук, они формируют сначала протофибриллы волоса, затем – микрофибриллы и, наконец, самые крупные волокна – макрофибриллы. Обвиваясь друг вокруг друга, макрофибриллы формируют основные волокна коркового слоя (рис.4). Фибриллы ориентированы параллельно оси волоса и параллельно друг другу. Они связываются между собой водородными и дисульфидными связями, которые образуют поперечные мостики между фибриллами. Если бы не дисульфидные мостики между остатками цистеина соседних белковых цепей, то цепи разошлись бы, и волокно распалось. Именно поперечные дисульфидные связи между остатками цистеина придают кератину его уникальные качества: прочность и эластичность. Но дисульфидные связи не очень прочные. В водном растворе дисульфидные мостики могут разъединяться и соединяться в новом положении. Например, если волос намочить и накрутить на бигуди, то перегруппировка дисульфидных связей приведет к тому, что высохший волос на какое-то время будет сохранять форму бигуди, – образуется завиток. На разрушении дисульфидных связей и формировании новых основан принцип химической завивки волос. Так как кортекс волоса, который главным образом обеспечивает прочность волоса, весь держится на дисульфидных связях, ему нужна дополнительная защита, в частности от воды. Функцию защиты волос от воды и механических воздействий выполняет барьерный слой – кутикула, которая также состоит из кератина, только более прочного.
Рисунок 4. Строение стержня волоса. Условные обозначения: 1-кутикула; 2- кортекс; 3- медуллярный слой
Кутикула представляет собой шесть-десять перекрывающихся слоев прозрачных черепицеобразно наложенных друг на друга плоских ороговевших клеток (кератиновых чешуек), содержащих в основном аморфный кератин. В кератине кутикулы очень много цистеина. Пространство между чешуйками кутикулы и между кутикулой и кортексом заполнено липидными пластами, напоминающими липидные пласты рогового слоя. Однако в волосе эта прослойка организована иначе. Если в роговом слое главную роль играют церамиды, то в волосах на первый план выступают жирные кислоты. В основном это разветвленная 18-углеродная метилэйкозаноевая кислота, которая связывается с цистеином тиоэфирной связью. Так как цистеина в кутикуле много, метилэйкозаноевая и другие жирные кислоты покрывают чешуйки кутикулы сплошным слоем (F-слой, или слой жирных кислот – fatty acid layer). Анализ липидного состава волос показывает, что в состав интегральных (ковалентно связанных с цистеином) липидов входит примерно 50% жирных кислот (из них 40% метилэйкозаноевой кислоты), 40% сульфата холестерина, 7% холестерина и 3% жирных спиртов. В составе полярных липидов (формирующих межклеточные липидные пласты) обнаружено около 60% церамидов, 7–10% гликофинголипидов, 30% сульфата холестерина.
Благодаря липидной прослойке и наличию дисульфидных связей, чешуйки кутикулы волоса плотно прилегают друг к другу. Так как все они располагаются в одной плоскости, луч света, падающий на волос, равномерно отражается от его поверхности – волос блестит. Поэтому блеск – обязательный признак здоровых волос. Среди средств, которые наносятся на волосы, присутствует много веществ, которые либо разрушают липидный слой (например, поверхностно–активные вещества шампуней), либо дезорганизуют дисульфидные связи. Это приводит к ослаблению связи между чешуйками кутикулы. В результате чешуйки кутикулы приподнимаются, зазубриваются и приходят в беспорядок. Свет, падая на такие волосы, отражается хаотично, и волосы перестают блестеть. Кроме того, зазубренные чешуйки кутикулы соседних волос цепляются друг за друга и слущиваются при расчесывании, обнажая кортекс, в результате волос теряет прочность и легко обламывается.
Заключение
Рассмотрев строение волоса, мы можем ответить на многие вопросы, относительно правильного выбора ингредиентов для средств по уходу за волосами, руководствуясь основным правилом – «не навреди». Следует учитывать, что волосы довольно активно реагируют на внешние воздействия. Например, при частом окрашивании и химической завивке, злоупотреблении термическими методами укладки волосы могут терять большой процент влаги и липидов. Наиболее разрушительно на структуру волос воздействует УФ – излучение, способствуя вымыванию из кутикулы цистеин-содержащего белка, который обеспечивает сцепление чешуек кутикулы, и разрушая липидную прослойку волос. Это означает, что для защиты волос необходимо использовать УФ – фильтры и антиоксиданты. Чтобы при мытье головы волосы не теряли необходимые для них липиды, следует выбирать мягкие системы ПАВ.
Поврежденные волосы нельзя «починить», но можно улучшить их внешний вид и предохранить от дальнейшего повреждения, используя кондиционирующие добавки, которые приглаживают чешуйки кутикулы. В качестве таких добавок можно использовать белки (кератин, протеины шелка, пшеницы и т.п.), липиды (церамиды), полисахариды (хитозан, гиалуриновая кислота), а также любые другие макромолекулы, способные закрепляться на волосах, не утяжеляя их. Для того чтобы кутикула вновь отрастающих волос была крепче и устойчивее к вредным воздействиям, используют масла, восстанавливающие липидный барьер волос – масла авокадо, репейное, аргановое, жожоба и др. Кондиционирующие добавки также эффективны для укрепления волос. При выборе добавок для борьбы с выпадением волос, следует учитывать, что основной мишенью в этом случае являются клетки волосяных фолликулов, а также сальные железы и кожа волосистой части головы. Здесь очень важным является время воздействия, поэтому лучше включать такого рода активы в несмываемые средства для волос. Поскольку при андрогенной алопеции происходит прогрессирующее истончение волос, то применение кондиционирующих добавок также является оправданным.
Строение и структура волос — салоны красоты «Kawaicat»
Инопланетная структура или нет?
В большинстве случаев мы не задумываемся о строении волос… Зато примерно «знаем» как покрасить волосы, какой рецепт наша бабушка в молодости использовала для укрепления и ускорен роста волос, ну или же наизусть знаем номер салона, где ежемесячно спасают наши волосы.
А порой нужна знать не так уж много «внутреннем мире» наших красивых и шелковистых спутников жизни, чтобы сохранить их качество.
Пришло время узнать, понять, каковы же они на вкус, эти волосы?)
Начнём с основ!
Строение волос
Абсолютно каждый человеческий волос на 80% состоит из белка-кератина, от 10% до 15% «жилплощади» занимает вода, ну а все остальное, примерно 5% — 10%,-пигменты, минералы и липиды.
Волос состоит из 3 частей:
1. Медула;
2. Кутикула
3. Кортепкс
Теперь нужно познакомится с каждым отдельно.
Первой составляющей является
Медула – это стержень волоса, который состоит из 2-4 слоев, не ороговевших клеток кубической формы. Медула отвечает за терморегуляцию, объем и силу волос . В наше время медула у многих прерывистая или отсутствует вообще. Раньше, когда ещё ниши предки танцевали вокруг костра и охотились на диких кабанов в шкурах тигров, волосы согревали в холодные времена и защищали от жары. Например, у индейцев медула была очень прочной.
Второй составляющей является
Кутикула – верхний слой волоса, который состоит из 6-10 слоев, удлиненных ороговевших пластинообразных клеток, которые по типу черепицы накладываются друг на друга по направлению от корней к концам. Кутикула прежде всего выполняет функцию воина, который яро сражается за безопасность и «спокойный сон» волоса. Именно этот слой отвечает за блеск и шелковистость волоса.
И последний не по значению, конечно же,
Кортекс – центральная, самая значимая по размерам часть волоса. Именно в кортексе происходят все химические процессы: окрашивание, химическая завивка и.т.д.
Кортекс или кора в свою очередь состоит из 2-х частей:
кератин — белковое вещество, именно белок, а точнее белковые нити обеспечивают прочность волос. Волосы почти на 90% состоят кератина.
Кератин построен в виде спирали. Отдельные цепочки кератина скреплены поперечно многочисленными связями (дисульфидными мостиками), что придает им дополнительную прочность. Дисульфидные мостики играют важную роль в химической завивке и выпрямлении.
При химической завивке волосам придают необходимую форму (с помощью коклюшек) разрушая дисульфидные мостики, а затем волосы подсушивают феном, при этом образуются новые мостики, которые фиксируются, что позволяет сохранить новую форму прически на длительное время.
Правда, в результате химической обработки волоса: окрашивания, блондирования, химической завивки и т.п., количество кератина резко снижается, что может привести к различного рода проблемам.
меланин — натуральный пигмент волоса, он содержит два пигмента:
а) эу–меланин,
б) фео–меланин,
помимо этого в волосе присутствуют мельчайшие пузырьки воздуха.
Цвет волос зависит от количества того или иного пигмента, а также от количества воздуха, который «разбавляет» пигмент.
Чем больше эу–меланина в волосах, тем темнее цвет волос и, наоборот, в светлых волосах эу–меланин практически отсутствует, на его месте преобладают пузырьки воздуха.
Например, седые волосы вместо пигмента содержат молекулы кислорода, поэтому их тяжелее прокрасить, чем другие волосы.
Если рассмотреть молекулы эу–меланина под микроскопом, мы увидим три основных цвета: синий, красный, желтый.
Когда мы осветляем волосы, в первую очередь растворяются синие молекулы, они отвечают за глубину и холодность тона. Это самые слабые молекулы. Именно пепельные оттенки быстрее всех вымываются из волоса, так как в основе содержат синий пигмент.
Красные молекулы отвечают за яркость и насыщенность цвета.
Желтые — самые устойчивые, осветляются плохо, отвечают за яркость цвета.
*именно поэтому, когда нас осветляют, мы похожи на цыплят)))
Фео–меланин, молекула маленького размера, округлой формы желтого и красного цвета. Фео–меланин рассеянный пигмент, более мелкий, он отвечает за фон осветления.
Молекулу фео–меланина мы не разрушаем, а осветляем до определенного уровня и проводим нейтрализацию нежелательных оттенков. Если мы разрушим фео-меланин, то разрушим структуру волоса.
«Немного много» информации о структуре волос, но зато теперь можно с уверенностью сказать, что волос — структура вполне земная, которая требует к себе чуткого внимания и бережного отношения! Берегите Ваши волосы, тем более теперь Вы имеете представление о внутреннем мире шелковистых и блестящих)
Ждём всех за подбором ухода и бережным окрашиванием у нас в студии каждый день с 11 до 21 по адресу улица Бауманская дом 11 строение 8
Все подробности так же по телефону
8(495)134-22-54
Формирование и функция кутикулы растений
Abstract
Кутикула растений представляет собой внеклеточный гидрофобный слой, покрывающий надземный эпидермис всех наземных растений, обеспечивая защиту от высыхания и внешних воздействий окружающей среды. За последнее десятилетие был отмечен значительный прогресс в сборке моделей для биосинтеза двух его основных компонентов, полимерного кутина и кутикулярных восков. Совсем недавно два прорыва в долгожданных молекулярных основах образования алканов и синтеза полиэфиров позволили построить почти полные пути биосинтеза как восков, так и кутина.Одновременно возникает сложная регуляторная сеть, контролирующая синтез кутикулы. Также стало ясно, что физиологическая роль кутикулы выходит далеко за рамки ее основной функции в качестве транспирационного барьера, играя важную роль в различных процессах, от развития до взаимодействия с микробами. Здесь мы рассматриваем недавний прогресс в биохимии и молекулярной биологии синтеза и функции кутикулы и выделяем некоторые из основных вопросов, которые будут определять будущие исследования в этой области.
Первые поселенцы растений на суше примерно 450 миллионов лет назад в среднюю палеозойскую эру столкнулись с устрашающим набором проблем, связанных с их новой земной средой, включая высыхание, экстремальные температуры, гравитацию и повышенное воздействие ультрафиолетового излучения (водные ресурсы). , 2003; Leliaert et al., 2011). Следовательно, переход от исключительно водного к наземному образу жизни потребовал бы эволюции набора морфологических и физиологических особенностей, некоторые из которых стали очевидными при изучении летописи окаменелостей или изучении существующих линий растений.Например, развитие архитектурно сложных клеточных стенок для биомеханической поддержки и структурной защиты, которые типичны для современных наземных растений, можно проследить до дивергенции и радиации внутри зеленых водорослей Charophycean, их непосредственных предков (Sørensen et al., 2011). Однако наиболее важным адаптивным признаком для выживания во время земледелия была способность удерживать воду во все более обезвоживающейся среде обитания. Следовательно, способность синтезировать, откладывать и поддерживать гидрофобный поверхностный слой или кутикулу на поверхности надземных органов, возможно, была одним из самых важных нововведений в истории эволюции растений.Эта идея подтверждается как свидетельствами окаменелостей (Edwards, 1993), так и повсеместностью кутикулы среди всех существующих эмбриофитов, от мохообразных (Budke et al., 2012) до покрытосеменных.
Обладая защитной кожей, вместе с рядом адаптивных стратегий для получения и сохранения воды, а также для предотвращения или выдерживания водного стресса, эмбриофиты теперь процветают в широком диапазоне высыхающих сред (Ogburn and Edwards, 2010; Aroca et al. al., 2012; Delaux et al., 2012; Jones, Dolan, 2012; Obata, Fernie, 2012; Gaff, Oliver, 2013).Соответственно, кутикула широкого круга видов и в различных экологических и сельскохозяйственных контекстах изучалась с точки зрения их роли в качестве основного барьера на пути транспирационной потери воды. Однако теперь ясно, что кутикула играет множество других ролей в развитии растений, физиологии и взаимодействии с абиотической средой и другими организмами. Действительно, в последние годы было много случаев неожиданных ассоциаций между кутикулой и различными аспектами биологии растений.Параллельно за последнее десятилетие наблюдался значительный прогресс в понимании биосинтеза основных компонентов кутикулы и сложных регуляторных сетей, которые контролируют синтез и сборку кутикулы.
Этот обзор суммирует недавний прогресс в выяснении биохимии и молекулярной биологии синтеза и функции кутикулы и подчеркивает некоторые связи с другими аспектами биологии растений, включая передачу сигналов, защиту от патогенов и развитие. Учитывая широкие возможности и ограниченное пространство, не все аспекты биосинтеза кутикулы рассмотрены подробно, и недавние специализированные обзоры, посвященные биомеханическим свойствам кутикулы (Domínguez et al., 2011), защитные функции (Reina-Pinto, Yephremov, 2009) и свойства транспортного барьера (Burghardt and Riederer, 2006) могут представлять дополнительный интерес. Кроме того, обсуждаются ключевые текущие вопросы в этой области и предлагаются возможные будущие подходы к их решению.
СТРУКТУРА КУТИКУЛЫ, БИОСИНТЕЗ И СБОРКА
Кутикула растений — это композитные структуры, состоящие из ковалентно связанных макромолекулярных каркасов кутина и различных растворимых в органических растворителях липидов, которые вместе называются восками.Хотя кутикулу обычно считают независимо от лежащей в основе полисахаридной клеточной стенки эпидермиса, эти две структуры физически связаны и имеют некоторые перекрывающиеся функции. Действительно, кутикулу можно рассматривать как специализированную липидную модификацию клеточной стенки, точно так же, как лигнификация является обычной модификацией вторичных клеточных стенок растений. Микроскопическая структура кутикулы часто делится на два домена на основе гистохимического окрашивания и их предполагаемого химического состава: богатый кутином домен со встроенными полисахаридами, который называется «кутикулярным слоем», и нижележащий слой, который менее многочисленен. в полисахаридах, но обогащенных парафином, называемых «собственно кутикулой» ().Воски либо откладываются внутри кутинной матрицы (внутрикутикулярный воск), либо накапливаются на ее поверхности в виде кристаллов или пленок эпикутикулярного воска. Эти эпикутикулярные воски могут придавать различные макроскопические свойства поверхности: эпикутикулярные пленки отвечают за глянцевый внешний вид, характерный для многих листьев и плодов, в то время как эпикутикулярные воски вызывают тусклый, сизый оттенок листьев брокколи ( Brassica oleracea ) и арабидопсиса ( Arabidopsis). thaliana ) стебли.Архитектурную организацию кутикулы можно определить с помощью ряда микроскопических методов. Сканирующая электронная микроскопия может выявить сложные и разнообразные морфологии эпикутикулярных кристаллов воска (; Jeffree, 2006), в то время как просвечивающая электронная микроскопия показывает отчетливую структуру внутренних слоев кутикулы, хотя этот подход не позволяет визуализировать восковые структуры (). Кутикулы значительно различаются по своей архитектуре и, в зависимости от вида и онтогенеза, резко различаются по толщине, от нанометра до микрометра (Jeffree, 2006).В последнем случае световая микроскопия может использоваться для выяснения тонких структур кутикулы и стенки эпидермальных клеток (), в то время как гистохимическое окрашивание в сочетании с конфокальной микроскопией может дополнительно разрешить трехмерную архитектуру кутикулы (Buda et al., 2009).
Строение кутикулы растений. A, схематическая диаграмма, показывающая основные структурные особенности кутикулы и нижележащего слоя эпидермальных клеток (не в масштабе). B. Изображение эпидермиса листа Arabidopsis и вышележащей кутикулы, полученное с помощью сканирующей электронной микрофотографии, в поперечном сечении.Бар = 5 мкм. (Изображение любезно предоставлено доктором Лейси Сэмюэлс.) C. Просвечивающая электронная микрофотография стенки и кутикулы стволовых эпидермальных клеток Arabidopsis. Бар = 500 нм. (Изображение любезно предоставлено доктором Кристиан Наврат.) D. Изображение световой микроскопии, показывающее кутикулу зрелого плода томата зеленой стадии, окрашенного Суданом красным, и клеточные стенки полисахарида, окрашенные альциановым синим. Бар = 50 мкм.
Биосинтез воска
Состав воска может существенно различаться в зависимости от вида, онтогенеза и условий роста в окружающей среде (Jenks and Ashworth, 1999).В большинстве случаев большинство соединений, составляющих кутикулярный воск, получают из жирных кислот с очень длинной цепью (ЖКОДЦ; C20 – C34), включая алканы, альдегиды, первичные и вторичные спирты, кетоны и сложные эфиры (). У некоторых видов различные липофильные вторичные метаболиты, такие как пентациклические тритерпеноиды, флавоноиды и токоферолы, также могут быть существенными компонентами (Jetter et al., 2006). Достигнут впечатляющий прогресс в раскрытии молекулярной биологии, лежащей в основе биосинтеза воска на основе ЖКОДЦ, и с этой целью Arabidopsis предоставил отличную экспериментальную модель (Bernard and Joubès, 2013).В дополнение к его хорошо известным преимуществам в качестве генетической системы, наличие эпикутикулярных кристаллов стебля воска, которые придают сизый цвет дикому типу, позволило легко провести скрининг мутантов с дефицитом воска. Такие мутанты, получившие название eceriferum ( cer ; Koornneef et al., 1989), обычно демонстрируют фенотип глянцевого стебля, и в первую очередь молекулярный анализ этих и других мутантов парафина показал, что путь биосинтеза ацильного парафина становится все более полным. был установлен.
Таблица I.
Основные классы ацил-липидов, обнаруженные в кутикулярных воскахБольшинство классов встречается в виде гомологических рядов с широким распределением длин цепей, показаны соединения с типичной средней длиной цепи.
Биосинтез воска начинается с биосинтеза жирных кислот de novo C16 или C18 в пластиде эпидермальных клеток (). Эти соединения длинноцепочечных жирных кислот превращаются в тиоэфиры CoA с помощью изофермента длинноцепочечной ацил-кофермента А-синтазы (LACS) и в конечном итоге переносятся в эндоплазматический ретикулум (ER).Механизм внутриклеточного транспорта жирных кислот из хлоропласта в ЭПР остается неизвестным, хотя гетерологичная экспрессия LACS1, LACS2 и LACS3 Arabidopsis облегчает поглощение жирных кислот дрожжами, что позволяет предположить, что этот класс ферментов может играть двойную роль в транспортировке жирных кислот и активации (Pulsifer et al., 2012). Для справки приводится список соответствующих генов, а также других генов, обсуждаемых в этом обзоре. C16-ацил-КоА тогда является субстратом для комплекса элонгазы жирных кислот (FAE).Благодаря последовательному добавлению двух атомов углерода за цикл, полученных из малонил-КоА, конечными продуктами этого комплекса являются ЖКОДЦ. Комплекс состоит из четырех основных субъединиц: β-кетоацил-КоА-синтазы, β-кетоацил-КоА-редуктазы, β-гидроксиацил-КоА-дегидратазы и еноил-КоА-редуктазы. У Arabidopsis 21 ген, как предполагается, кодирует β-кетоацил-CoA-синтазу, а для биосинтеза воска наиболее важным геном, основанным на мутантном фенотипе, является CER6 (Fiebig et al., 2000). Гены, кодирующие остальные субъединицы комплекса FAE, представленные KCR1 , PAS2 и CER10 , соответственно, менее избыточны, а их плейотропные мутантные фенотипы подчеркивают общую важность FAE в генерации предшественников VLCFA для биосинтеза сфинголипидов. (Чжэн и др., 2005; Бах и др., 2008; Beaudoin et al., 2009). Дополнительное семейство белков, состоящее из CER2, CER26 и CER26-подобных, по-видимому, необходимо для удлинения жирных кислот до длин, превышающих 28C (Haslam et al., 2012; Pascal et al., 2013). Любопытно, что эти ферменты имеют гомологию последовательностей с ацилтрансферазами BAHD, но консервативные каталитические аминокислотные остатки этого семейства ферментов незаменимы для стимулирующей удлинение активности CER2 (Haslam et al., 2012). Циклы элонгации могут быть прерваны тиоэстеразой с образованием свободных VLCFA, или сложные эфиры VLCFA-CoA могут подвергаться дальнейшим модификациям.
Пути биосинтеза кутина и воска. Гены (синий текст) описаны в обзоре. Красный текст обозначает классы соединений, которые обычно наблюдаются в кутикулярных восковых смесях.
Таблица II.
Гены, связанные с кутикулой, обсуждаемые в этом обзоре
| Символ гена | Название гена | Виды | Идентификатор локуса | Описание | ||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ABCG11 | ATP-BINDING CASSETTE G | Arabidopsis | At1G17840 | Полутранспортер ABC | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ABCG13 | ATP-BINDING CASSETTE G13 | Arabidopsis | At1G51460 | ABC16 | Полутранспортер ABC -СВЯЗИВАЮЩАЯ КАССЕТА G32 | Arabidopsis | At2G26910 | Полный транспортер ABC | ||||||||||||||||||||||||||||||||||||||||||||||||
| BDG | BODYGUARD | Arabidopsis | At1G64670 | Семейство белков α / β-G64670 | B | BODYGUARD3 | Arabidopsis | At4G24140 | Гомолог BDG | |||||||||||||||||||||||||||||||||||||||||||||||
| CD1 | CUTIN DEFICIENT1 | Tomato | Solyc11G0062507 Cutin DEFICIENT1 | CUTIN DEFICIENT2 | Tomato | Solyc01G0 | Гомеодомен-лей молния IV фактор транскрипции | | |||||||||||||||||||||||||||||||||||||||||||||||||
| CER1 | ECERIFERUM1 | Инвазивный арабидопсис | в 1 | ECERIFERUM10 | Arabidopsis | At3G55360 | Enoyl-CoA редуктаза | |||||||||||||||||||||||||||||||||||||||||||||||||
| CER2 | ECERIFERUM2 | Arabidopsis | R510 требуется для удлинения жирных кислот от C28 до C30 | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| CER26 | ECERIFERUM26 | Arabidopsis | At4G13840 | Гомолог CER2, необходимый для удлинения жирных кислот выше C30 | ECERIFERUM26-подобный | Arabidopsis | At3G23840 | Гомолог CER2 и CER26 | ||||||||||||||||||||||||||||||||||||||||||||||||
| CER3 | ECERIFERUM3 | Образование арабидопсиса | At5G75||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ECERIFERUM1 | Arabidopsis | At4G33790 | VLCFA-CoA жирным ацил-CoA | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| CER5 / ABCG12 | ECERIFERUM5 / ATP-BINDING ATP-BINDING | ABC полуприцеп r | | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CER6 | ECERIFERUM6 | Arabidopsis | At1G68530 | β-кетоацил-КоА-синтаза | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| CER7 | ECERIFERUM7 900GISSO | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CER9 | ECERIFERUM9 | Arabidopsis | At4G34100 | Предполагаемая убиквитин-лигаза E3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| CFL1 | CURLY FLAG LEAF16 | RURLY FLAG LEAF16домен RURLY FLAG LEAF1 | , содержащий домен Osice2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| CYP77A6 | CYP77A6 | Arabidopsis | At3G10570 | CYP77A подсемейство цитохрома P450 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| CYP86A4 | CYP86A4 | CYP86A4 1600 | CYP86A подсемейство цитохрома P450 | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| DCR | ДЕФЕКТ В КУЛИКУЛЯРНЫХ РИДЖАХ | Arabidopsis | At5G23940 | BAHD ацилтрансфераза FD74 FD74 | Arabid 49 AD At2G26250 | β-кетоацил-КоА-синтаза | ||||||||||||||||||||||||||||||||||||||||||||||||||
| GPAT6 | ГЛИЦЕРОЛ-3-ФОСФАТ SN-2-АЦИЛТРАНСФЕРАЗА6 | Арабидопсис | -3-2G38110 | глицерол-бифункция ацилтрансфераза / фосфатаза|||||||||||||||||||||||||||||||||||||||||||||||||||||
| HDG1 | HOMEODOMAIN GLABROUS1 | Arabidopsis | At3G61150 | Homeodomain-Leu zipper IV фактор транскрипции | HTH16 HTH16 | HTH 16 4| Белок семейства Glc-метанол-холин оксидоредуктазы | | |||||||||||||||||||||||||||||||||||||||||||||||||
| IRG1 | ИНГИБИТОР ДИФФЕРЕНЦИАЦИИ ЗЕРНОВЫХ ПРОБК 1 | M.truncatula | Medtr5G014400 | C 2 H 2 Фактор транскрипции цинкового пальца | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| KCR1 | β-KETOACYL-COENZYME A REDUCTASE1 | 75 Arabidopsis редуктаза|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LACS1 | ДЛИННОЦЕПНАЯ СИНТАЗА АЦИЛ-КОЭНЗИМА A1 | Арабидопсис | At2G47240 | Длинноцепочечная ацил-КоА-синтаза | ACYL-CoA-синтазаACYL-CoA-синтаза LACS2 LACS COENZYME A SYNTHASE2 | Arabidopsis | At1G49430 | Длинноцепочечная ацил-CoA-синтаза | ||||||||||||||||||||||||||||||||||||||||||||||||
| LACS3 | ДЛИННАЯ ЦЕПЬ ACYL-COENZYME A SYNTHASE3 74 | Acyl-COENZYME A SYNTHASE3 74 | -CoA-синтаза||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LCR | LACERATA | Arabidopsis | At2G45970 | CYP86A подсемейство цитохрома P450 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| LTL1 | LI-TOLERANT LIPASE1 | Arabidopsis | CDGPolog -ANCHORED LIPID TRANSFER PROTEIN1 | Arabidopsis | At1G27950 | GPI-заякоренный липидный переносящий белок | ||||||||||||||||||||||||||||||||||||||||||||||||||
| LTPG2 | GPI-ANCHORED LIPID TRANSFER PROTEIN2 | Arabid липидный перенос | Arabid липидный переносчик | белок | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| MAh2 | MIDCHAIN ALKANE HYDROXYLASE1 | Arabidopsis | At1G57750 | CYP96A подсемейство цитохрома P450 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| MyB106 is | At3G01140 | Фактор транскрипции Myb | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MYB16 | Myb ДОМЕННЫЙ БЕЛК16 | Arabidopsis | At5G15310 | Фактор транскрипции Myb | DOMAIN MYB30 ArabidopsisAt3G28910 | Фактор транскрипции Myb | ||||||||||||||||||||||||||||||||||||||||||||||||||
| MYB41 | Myb ДОМЕННЫЙ БЕЛК41 | Arabidopsis | At4G28110 | Фактор транскрипции Myb | Myb DOMAIN | |||||||||||||||||||||||||||||||||||||||||||||||||||
| At5G62470 | Фактор транскрипции Myb | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OCL1 | ВНЕШНИЙ КЛЕТОЧНЫЙ СЛОЙ 1 | Кукуруза | GRMZM2G026643 | Фактор транскрипции Homeodomain-Leu zipper IV | PAS2 | PASTICCINO2 | Arabidopsis | At5G10480 | β-гидроксиацил-КоА-дегидратаза | |||||||||||||||||||||||||||||||||||||||||||||||
| SHN2 | SHINE2 | At5G10480 SHINE2 | At5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| WIN1 / SHN1 | WAX INDUCER1 / SHINE1 | Arabidopsis | At1G15360 | AP2-домен, содержащий фактор транскрипции | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| WSD1 | WAXLASE AYLACYLASYLACYLACYLACIACYLACYLACYLACYLACIACYLACI | Arabidopsis | At5G37300 | Воск-синтаза / ацил-КоА: белок семейства диацилглицерин-ацилтрансфераз | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| WXP1 | WAX PRODUCTION1 | M.truncatula | Medtr5G062700 | Фактор транскрипции, содержащий домен AP2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| — | — | Arabidopsis | At5G33370 | Гомолог CD1 |
жирные ацил-A КоА-редуктаза, фермент, кодируемый CER4 у Arabidopsis (Rowland et al., 2006). В восковой смеси могут присутствовать свободные первичные спирты, или они могут быть этерифицированы до жирной кислоты с образованием сложных эфиров парафина.В этом случае спирт связан с ацильной группой, производной от жирного ацил-КоА. Фермент Arabidopsis, ответственный за это, — WSD1, фермент семейства воскосинтазы / диацилглицерин ацилтрансферазы (Li et al., 2008).
Вторая ветвь биосинтеза ацильного воска приводит к образованию альдегидов и, в конечном итоге, алканов. Интересно, что у арабидопсиса LACS1, который также необходим для биосинтеза мономера кутина C16, по-видимому, обладает дополнительной специфичностью к C30 VLCFA и необходим для нормального накопления последующих восковых соединений (Lü et al., 2009). Это предполагает, что преобразование внутриклеточного пула свободных VLCFA обратно в VLCFA-CoA является важным путем для биосинтеза альдегидов и алканов, а не VLCFA-CoA, непосредственно полученных из FAE. В биосинтезе парафина давно не решен вопрос о ферментативной основе синтеза алканов. Классическая биохимия с использованием сырых экстрактов гороха ( Pisum sativum ) показала, что реакция, вероятно, происходит через восстановление VLCFA-CoA до промежуточного альдегида с последующим декарбонилированием с образованием алкана, который на 1 ° C короче (Cheesbrough and Kolattukudy, 1984; Шнайдер-Бельхаддад и Колаттукуди, 2000).Хотя этот фермент не был очищен и идентифицирован, недавно в результате исследований Arabidopsis были получены убедительные доказательства того, что CER1 и CER3 в комплексе действуют вместе, катализируя образование алканов из VLCFA-CoA. С помощью двухгибридного анализа расщепленного убиквитина дрожжей и анализа расщепленной люциферазы Arabidopsis было показано, что CER1 взаимодействует с CER3, а также с несколькими изоформами цитохрома b 5 . Кроме того, гетерологичная экспрессия комбинации CER1, CER3, цитохрома b 5 и LACS1 в дрожжах приводила к образованию алканов с очень длинной цепью (Bernard et al., 2012). Это убедительно свидетельствует о том, что комплекс, включающий CER1 и CER3 с цитохромом b 5 в качестве донора электронов, катализирует восстановление и декарбонилирование VLCFA-CoA с образованием кутикулярных алканов. Помимо того, что они являются основным компонентом смеси парафинов, алканы могут подвергаться дальнейшей модификации с образованием вторичных спиртов и кетонов. У Arabidopsis оба этих окисления осуществляются ферментом цитохрома P450 алкангидроксилазой средней цепи (MAh2; Greer et al., 2007).
Синтез предшественников кутина
Кутин обычно состоит из переэтерифицированных гидроксижирных кислот с меньшими количествами глицерина, фенилпропаноидов и дикарбоновых кислот (Kolattukudy, 2001). Химические процессы, которые расщепляют сложноэфирные связи, такие как омыление, легко высвобождают эти мономерные компоненты, хотя у некоторых видов дополнительный липидный полимер, называемый кутаном, остается устойчивым к такой обработке. Кутан богат эфиром и связями C-C, но его структура в остальном неизвестна, и, по-видимому, он ограничен относительно небольшим количеством существующих видов (Gupta et al., 2006). Гидроксижирные кислоты кутина обычно представляют собой ω-гидроксижирные кислоты, обычно с одной или двумя дополнительными гидроксильными группами средней цепи или эпоксидной группой (). Несмотря на обширные исследования химического состава кутинов растений в 1970-х и 1980-х годах (Kolattukudy, 2001), состав кутина Arabidopsis не был определен до относительно недавнего времени (Bonaventure et al., 2004; Franke et al., 2005). Важно отметить, что у этого важного модельного вида кутин стеблей и листьев нетипичен в том смысле, что его основным компонентом является дикарбоновая кислота (), подразумевая, что преобладающим структурным мотивом должен быть сополимер с неизвестным полигидроксисоединением, предположительно глицерин (Pollard et al., 2008). Однако, несмотря на атипичный состав кутина, арабидопсис оказался важной моделью для расшифровки пути биосинтеза кутина, а недавно было обнаружено, что кутин его цветочных органов более типичен, поскольку он состоит в основном из 10,16-дигидроксигексадекановой кислоты (Li-Beisson et al., 2009).
Типичные кутиновые мономеры и полимерная структура. A, Некоторые типичные мономеры кутина, полученные из жирных кислот C16 и C18. Сверху вниз: 10,16-дигидроксигексадекановая кислота, 16-гидроксигексадекановая кислота, 9,10-эпоксиоктадекановая кислота, 9,10,18-тригидроксиоктадекановая кислота и октадека-цис-6, цис-9-диен-1,18- двояковыпуклый, главный кутиновый мономер стеблей и листьев Arabidopsis.B, линейные и разветвленные домены, которые стали возможными благодаря различным сложноэфирным связям 10,16-дигидроксигексадекановой кислоты, схематически изображены, как указано.
В то время как существует значительное разнообразие в структуре мономеров кутина, путь биосинтеза кутина на основе 10,16-дигидроксигексадекановой кислоты является наиболее полным, и основные темы биосинтеза кутина, вероятно, являются общими для других мономеров кутина. Здесь мы суммируем этот путь на основе недавних молекулярно-генетических и биохимических исследований с использованием арабидопсиса и томатов ( Solanum lycopersicum ).
Внутриклеточные ацилтрансферазы и гидроксилазы
Биосинтез кутина начинается с синтеза жирных кислот de novo в пластиде эпидермальных клеток (). Следующие три стадии происходят в ER и состоят из ω-гидроксилирования и гидроксилирования средней цепи, а также синтеза промежуточного соединения ацил-CoA. Относительный порядок этих стадий неизвестен, хотя было показано, что ω-гидроксилирование предшествует гидроксилированию средней цепи и что конечный продукт этих стадий, скорее всего, представляет собой сложный эфир дигидроксигексадекановой кислоты и CoA (Li-Beisson et al., 2009). Ω-гидроксилаза кодируется членами подсемейства CYP86 цитохрома P450s ( CYP86A4 в цветках арабидопсиса; Li-Beisson et al., 2009), в то время как гидроксилаза средней цепи кодируется подсемейством CYP77 ( CYP77A6 в цветках арабидопсиса). ; Li-Beisson et al., 2009). Ацилтрансферазы, которые синтезируют ацил-КоА, кодируются семейством LACS, которое состоит из девяти членов у Arabidopsis, и как LACS1, , так и LACS2 , по-видимому, ответственны за биосинтез мономера кутина C16 (Lü et al., 2009).
Дополнительная внутриклеточная ацилтрансфераза, необходимая для синтеза полиэфира кутина, представляет собой глицерин-3-фосфатацилтрансфераза (GPAT). Недавно было показано, что растения обладают уникальным подсемейством бифункциональных GPAT, кодирующих ферменты, обладающие как sn-2 -специфической активностью глицерин-3-фосфат: ацил-КоА-ацилтрансферазы, так и активностью фосфатазы с образованием 2-моноацилглицерилового эфира (Yang и др., 2010). В случае цветочного кутина Arabidopsis эта активность кодируется GPAT6 (Li-Beisson et al., 2009). Хотя конкретная последовательность всех этапов внутриклеточного биосинтеза потребует дополнительной характеристики субстратной специфичности каждого фермента, биохимическая характеристика бифункциональных GPAT Arabidopsis указывает на то, что они имеют сильное предпочтение к ω-гидроксилированному ацил-КоА, предполагая, что гидроксилирование предшествует переносу на глицерин. (Ян и др., 2012). В любом случае конечным продуктом внутриклеточных стадий биосинтеза кутина, вероятно, будут 2-моноацилглицериловые эфиры мономеров кутина.В случае кутина на основе 10,16-дигидроксигексадекановой кислоты это 2-моно (10,16) -дигидроксигексадеканоилглицерин (2-MHG).
Транспорт предшественников кутикулы
После синтеза предшественников воска и кутина они экспортируются из ER через плазматическую мембрану, через полисахаридную клеточную стенку и к формирующейся кутикулярной мембране. Большинство этих транспортных процессов плохо изучены, хотя было показано, что транспортировка предшественников воска и кутина через плазматическую мембрану зависит от переносчиков АТФ-связывающих кассет (ABC).У Arabidopsis для экспорта парафина требуются CER5 / ABCG12 (Пигин и др., 2004) и ABCG11 (Берд и др., 2007). Оба они кодируют полутранспортеры, и на основании анализа двойных мутантов и анализов комплементации бимолекулярной флуоресценции было высказано предположение, что гетеродимер ABCG11 / ABCG12 необходим для секреции воска (McFarlane et al., 2010). ABCG11 также необходим для накопления кутина, и, поскольку он также способен димеризоваться сам с собой, было высказано предположение, что этот гомодимер является функциональным комплексом, ответственным за экспорт кутина (McFarlane et al., 2010). Кроме того, было показано, что третий полупереносчик Arabidopsis, ABCG13, необходим для отложения кутина в цветках (Panikashvili et al., 2011).
Совсем недавно полные переносчики, необходимые для отложения кутина, были идентифицированы у Arabidopsis (ABCG32; Bessire et al., 2011), а также у дикого ячменя ( Hordeum spontaneum ) и риса ( Oryza sativa ; Chen et al., 2011). ). Несмотря на четкие генетические доказательства, подтверждающие роль транспортеров ABC в экспорте липидов в кутикулу, субстратная специфичность этих транспортеров еще не была продемонстрирована in vitro.Однако все переносчики ABC, которые участвуют в биосинтезе кутикулы на сегодняшний день, являются членами подсемейства ABCG, которое связано с транспортом липидов и гидрофобных соединений в др. Системах (Moitra et al., 2011). Более того, в нескольких случаях внутриклеточные липидные включения наблюдались у мутантов переносчиков ABC, дополнительно подтверждая их прямое участие в экспорте кутикулярных липидов (Pighin et al., 2004; Bird et al., 2007; Bessire et al., 2011).
Экспорт некоторых парафиновых соединений, по-видимому, также облегчается заякоренными гликозилфосфатидилинозитолом (GPI) белками переноса липидов (LTP), LTPG1 и LTPG2, которые связаны с внеклеточной стороной плазматической мембраны (Debono et al., 2009; Ли и др., 2009; Ким и др., 2012). Эти белки представляют собой уникальный класс LTP, семейство небольших и обычно растворимых белков, которые связывают различные липидные субстраты in vitro (Yeats and Rose, 2008). Главный остающийся вопрос заключается в том, как гидрофобные предшественники кутикулы транспортируются через гидрофильную среду полисахаридной клеточной стенки к кутикуле. Предполагается, что апопластные LTP играют роль, хотя генетические или биохимические доказательства их участия в транспорте в целом отсутствуют (Yeats and Rose, 2008).В случае предшественника дигидроксиацилкутина 2-MHG, фрагмент глицерина придает достаточную полярность, чтобы обеспечить растворимость в воде при низких миллимолярных концентрациях (Yeats et al., 2012b). Это предполагает, что липид-связывающие белки или другие факторы не являются необходимыми для облегчения транспорта этого основного предшественника биосинтеза кутина. Однако растворимость глицериловых эфиров менее полярных мономеров кутина не исследовалась, и для них, наряду с восками, могут потребоваться дополнительные факторы для увеличения их растворимости в апопласте.
Полимеризация кутина
Заключительным этапом синтеза кутина является включение гидроксиацильного мономера в полимер, но молекулярный механизм полимеризации кутина был давней загадкой. Недавний прогресс в этой области был достигнут путем изучения мутанта кутина с дефицитом кутина1 ( cd1 ) и трансгенных растений томата, в которых экспрессия CD1 подавлялась с использованием стратегии РНК-интерференции (Girard et al., 2012; Yeats et al., 2012б). Мутант cd1 демонстрирует серьезное снижение количества полимеризованного кутина в кутикуле плода (Isaacson et al., 2009), хотя химический анализ показал, что, в отличие от плодов дикого типа, плоды мутанта накапливают неполимеризованный 2-MHG ( Yeats et al., 2012b). Клонирование мутированного гена показало, что он кодирует белок семейства липаз / гидролаз (GDSL) с мотивом GDSL, который локализуется в развивающейся кутикуле (Girard et al., 2012; Yeats et al., 2012b). Несмотря на сходство с липолитическими ферментами, рекомбинантный белок действует как ацилтрансфераза in vitro, образуя полиэфирные олигомеры из 2-MHG (Yeats et al., 2012б).
Идентификация CD1 как первой известной кутинсинтазы поднимает несколько вопросов о специфичности и общности реакции, которую он катализирует. Филогенетический анализ CD1 и гомологичных генов показывает, что, несмотря на принадлежность к очень большому семейству генов, подсемейство GDSL, представленное CD1 , относительно невелико и хорошо консервативно, с последовательностями, представленными в различных таксонах наземных растений (Volokita et al. , 2011). У арабидопсиса его предполагаемые ортологи образуют семейство генов, состоящее из пяти членов, и подавление экспрессии двух из них ( LTL1 и At5g33370 ) привело к тому, что растения демонстрируют слияние органов цветков и не имеют наноразмеров на поверхности лепестков, фенотипы которых являются соответствует дефициту кутина (Shi et al., 2011). Дополнительный предполагаемый ортолог CD1 из Agave americana обнаруживает сходную локализацию и экспрессию, дополнительно подтверждая консервативный механизм CD1-подобных ферментов, действующих как кутинсинтазы (Reina et al., 2007). Несмотря на присутствие нулевого аллеля, мутант cd1 и не полностью лишен кутина, поэтому идентичность дополнительных кутинсинтаз или, возможно, неферментативных механизмов синтеза кутина, представляет собой интригующее направление будущих исследований.
Полимерная структура кутина изучена недостаточно.Мономерная композиция может обеспечить «список частей», но относительное количество возможных связей в полимере трудно определить, в основном из-за сложности солюбилизации интактного кутина (Serra et al., 2012). Тем не менее, множественные функциональные возможности, присутствующие во многих мономерах кутина, указывают на то, что нативные полимеры кутина могут варьироваться от линейных до разветвленных или поперечно-сшитых структур (Pollard et al., 2008). Например, в идеализированном полимере кутина, состоящем исключительно из 10,16-дигидроксигексадекановой кислоты, мономеры могут быть соединены этерификацией либо концевой гидроксильной группы, либо гидроксильной группы средней цепи.Этерификация одного гидроксила приведет к линейному полимеру, тогда как этерификация обеих гидроксильных групп приведет к образованию разветвленных структур (). Идентификация гидроксильных групп, которые этерифицируются CD1 и другими кутинсинтазами, должна указывать на то, контролируется ли региоспецифичность полимеризации кутина ферментативно и катализируют ли специфические кутинсинтазы образование линейных или разветвленных доменов полимера кутина. Более того, неизвестно, как ветвление или перекрестное связывание кутина влияет на функцию кутикулы, и идентификация дополнительных кутин-синтаз позволит исследовать это с использованием генетических подходов.
РЕГУЛЯЦИЯ БИОСИНТЕЗА Кутикулы
Регуляция биосинтеза кутикулы является сложной и включает в себя взаимодействующие сигнальные сети, связанные с реакциями на стресс окружающей среды, ответами патогенов и регуляцией обратной связи, основанной на структуре и целостности самой кутикулы. Более того, поскольку кутикула синтезируется исключительно эпидермальными клетками, регуляция идентичности эпидермиса во время развития также может рассматриваться как играющая регулирующую роль в развитии кутикулы.Подробнее об этом можно прочитать в прекрасном обзоре Javelle et al. (2011), и мы сосредоточимся здесь только на прямых регуляторах биосинтеза кутина и воска (). Даже в этом ограниченном контексте анализ регуляторных мутантов осложняется компенсаторными механизмами между кутином и биосинтезом воска и другими плейотропными фенотипами. Тем не менее, начинает возникать сложная регуляторная сеть, которая реагирует на сигналы развития и окружающей среды, опосредованные гормонами, факторами транскрипции и посттранскрипционной регуляцией.
Регуляция биосинтеза кутикулы. Показано краткое изложение взаимодействия факторов окружающей среды и регуляторных генов, которые, как известно, влияют на биосинтез кутина или воска.
Окружающая среда и гормоны
Систематический анализ как состава кутикулы, так и экспрессии генов у Arabidopsis показывает, что синтез воска индуцируется нехваткой воды, хлоридом натрия и обработкой абсцизовой кислотой (ABA) (Kosma et al., 2009). Напротив, биосинтез кутина индуцировался только дефицитом воды, а не АБК или хлоридом натрия, что позволяет предположить, что, по крайней мере, у Arabidopsis обнаружение различных осмотических стрессов является сложным и лишь частично зависит от АБК (Kosma et al., 2009). Однако, учитывая, что АБК уже хорошо зарекомендовала себя в качестве регулятора реакции растений на дефицит воды посредством регулирования устьичной апертуры (Lee and Luan, 2012), регулирование биосинтеза кутикулы с помощью АБК является интересной областью для дальнейших исследований, направленных на понимание и инженерию засухи. толерантность в посевах.
Кроме того, было показано, что обработка в темноте и холодом снижает экспрессию некоторых компонентов комплекса FAE (Hooker et al., 2002; Joubès et al., 2008).Было показано, что несколько генов биосинтеза воска индуцируются бактериальными патогенами (Raffaele et al., 2008) и во время заражения пшеницы ( Triticum aestivum ) гессенской мухой ( Mayetiola destructor ; Kosma et al., 2010), но в целом значение индукции синтеза кутикулы для устойчивости к вредителям или патогенам изучено плохо.
Факторы транскрипции и биосинтез кутикулы
Первым геном фактора транскрипции, который, как было установлено, играет роль в регуляции биосинтеза кутикулы, был домен AP2 WAX INDUCER1 / SHINE1 ( WIN1 / SHN1 ; Aharoni et al., 2004; Broun et al., 2004). Сверхэкспрессия этого гена привела к появлению глянцевых листьев с большей восковой нагрузкой, чем у дикого типа, и более низкой транспирацией, хотя это, вероятно, было связано с уменьшенной плотностью устьиц, а не с восковым фенотипом (Aharoni et al., 2004). Более поздние исследования показали, что уровни кутина также увеличиваются у растений с избыточной экспрессией WIN1 / SHN1 и что повышенная регуляция генов, кодирующих ферменты биосинтеза кутина, предшествует индукции генов биосинтеза воска (Kannangara et al., 2007). WIN1 / SHN1 является частью трехчленного семейства генов у Arabidopsis, и подавление всех трех генов привело к снижению количества кутина, но не восков (Shi et al., 2011). Эти авт. Также продемонстрировали, что эти факторы транскрипции непосредственно активируют промоторы нескольких генов биосинтеза кутина, дополнительно подтверждая первичную роль в регуляции кутина с последующим действием на биосинтез воска (Shi et al., 2011). Помимо регуляции биосинтеза кутина, факторы транскрипции SHN также индуцировали экспрессию нескольких пектин-модифицирующих ферментов, что свидетельствует о координации синтеза кутикулы с полисахаридной клеточной стенкой (Shi et al., 2011). Эта вторая функция факторов транскрипции SHN в регуляции полисахаридной клеточной стенки дополнительно подтверждается экспериментами, в которых избыточная экспрессия SHN2 Arabidopsis в рисе привела к значительному увеличению количества целлюлозы и сопутствующему снижению лигнина (Ambavaram et al. , 2011). С другой стороны, на общую роль факторов транскрипции, связанных с WIN1 / SHN1, в регуляции синтеза кутина указывают исследования ортологичных генов ячменя ( Hordeum vulgare ; Taketa et al., 2008) и томат (Shi et al., 2013). Таким образом, совокупность данных свидетельствует о том, что факторы транскрипции SHN координируют не только синтез кутина, но и полисахаридную клеточную стенку эпидермиса. Это в конечном итоге подчеркивает тот факт, что кутикула представляет собой специализированную модификацию клеточной стенки и, подобно другим модификациям, таким как лигнификация или суберизация, ее следует рассматривать в контексте компонентов полисахаридной клеточной стенки. Помимо семейства SHN, другие факторы транскрипции домена AP2 из разных клад также могут играть роль в регуляции кутикулы.Например, сверхэкспрессия WXP1 из Medicago truncatula в люцерне ( Medicago sativa ) индуцировала продукцию воска (Zhang et al., 2005).
Недавно два фактора транскрипции, MYB106 и MYB16, были идентифицированы как регуляторы биосинтеза кутикулы, которые действуют аналогично WIN1 / SHN1 (Oshima et al., 2013). Оба они, по-видимому, действуют выше и непосредственно активируют WIN1 / SHN1 , но также и некоторые гены биосинтеза кутикулы (Oshima et al., 2013). Некоторые другие факторы транскрипции семейства MYB также участвуют в регуляции биосинтеза воска и кутина в ответ на стрессы окружающей среды. MYB30 индуцируется во время инфекции бактериальными патогенами, что приводит к активации нескольких генов комплекса FAE, а эктопическая сверхэкспрессия MYB30 ведет к увеличению восковой нагрузки (Raffaele et al., 2008). MYB96 был идентифицирован как АБК-индуцируемый фактор транскрипции, который обеспечивает устойчивость к засухе (Seo et al., 2009), отчасти из-за индукции биосинтеза воска в результате прямой активации MYB96 промоторов нескольких генов синтеза воска (Seo et al., 2011). В то время как MYB96 положительно регулирует выработку воска в ответ на стресс, MYB41 опосредует отрицательную регуляцию биосинтеза кутина в ответ на аналогичные стрессы. MYB41 индуцируется АБК, засухой и осмотическим стрессом, что ведет к подавлению генов биосинтеза кутина и нарушению структуры кутикулы (Cominelli et al., 2008).
Другой регуляторный фактор был идентифицирован посредством характеристики гена риса CURLY FLAG LEAF1 ( CFL1 ), который кодирует белок, содержащий домен WW, который негативно регулирует биосинтез кутикулы.Исследования ортологичного гена CFL1 у Arabidopsis показали, что он подавляет биосинтез кутина путем подавления функции HDG1, фактора транскрипции гомеодомена-лейциновой молнии IV (HD-ZIP IV), который, как было показано, индуцирует экспрессию нескольких гены биосинтеза кутина (Wu et al., 2011). Более общая роль белков HD-ZIP IV в регуляции синтеза кутина подтверждается также гомологичным геном томата CD2 , который необходим для биосинтеза кутина в плодах и других органах (Isaacson et al., 2009; Надакудути и др., 2012). Было показано, что у кукурузы ( Zea mays ) ген HD-ZIP IV OUTER CELL LAYER1 ( OCL1 ) является эпидермис-специфичным положительным регулятором биосинтеза воска, хотя кутин не определялся количественно у растений, сверхэкспрессирующих этот ген ( Javelle et al., 2010). Интересно, что белки HD-ZIP IV также участвуют в регуляции других специфичных для эпидермиса процессов, таких как дифференцировка трихомов и образование корневых волосков и замыкающих клеток устьиц (Masucci et al., 1996; Накамура и др., 2006; Takada et al., 2013). Учитывая их дополнительную связь с биосинтезом кутикулы, похоже, что общая черта членов этого семейства белков играет ключевую роль в биологии эпидермиса растений и в определении судьбы эпидермальных клеток.
Beyond Transcription Factors
В дополнение к сети факторов транскрипции, которые регулируют биосинтез кутикулы, недавно были обнаружены регуляторные механизмы, которые не включают прямую активацию транскрипции или репрессию за счет связывания промотора.Недавний пример явился результатом исследований мутанта Arabidopsis cer9 , который обнаруживает изменения в количестве и составе восков листьев и стеблей. Клонирование гена CER9 показало, что он кодирует белок со сходной последовательностью с дрожжевым Doa10, убиквитинлигазой E3, участвующей в ER-ассоциированной деградации неправильно свернутых белков (Lü et al., 2012). Учитывая локализацию в ER процессов биосинтеза воска и кутина, авт. Предположили роль CER9 в гомеостазе ключевых уровней ферментов биосинтеза кутикулы.Эксперименты, направленные на дальнейшее рассмотрение этой гипотезы, будут особенно интересными, учитывая неожиданное открытие, что мутант cer9 на самом деле демонстрирует повышенную засухоустойчивость и эффективность использования воды (Lü et al., 2012).
Один из наиболее интересных механизмов регуляции кутикулы возник в результате характеристики мутанта cer7 . CER7 кодирует экзосомальную экзорибонуклеазу, а мутант cer7 демонстрирует снижение содержания воска стебля и транскрипцию CER3 , основного фермента биосинтеза воска (Hooker et al., 2007). Недавно были идентифицированы два супрессора cer7 , которые восстанавливают транскрипт CER3 и уровни стебля воска, и клонирование соответствующих генов идентифицировало RDR1 и SGS3 , два консервативных компонента РНК-опосредованного пути молчания генов ( Лам и др., 2012). Была предложена модель, в которой CER7 участвует в деградации малых видов РНК, которые негативно регулируют транскрипт CER3. Будущая работа по идентификации таких малых видов РНК и других компонентов этого пути будет особенно интересной, поскольку ни один из известных видов малых РНК растений не картирован в CER7-зависимую область промотора CER3 (Lam et al., 2012).
ЭНИГМАТИЧЕСКИЕ ФАКТОРЫ В БИОСИНТЕЗЕ КУТИКУЛ
В дополнение к охарактеризованным компонентам биосинтеза кутикулы, которые могут быть включены в согласованную модель, как обсуждалось выше, были идентифицированы несколько генов / белков, которые необходимы для образования кутикулы, но не имеют четкого ассоциированного биохимическая функция, которая поместит их в определенную точку на пути. Одним из примеров является HOTHEAD (HTH), белок семейства Glc-метанол-холин оксидоредуктазы, который необходим для правильной организации кутикулы (Krolikowski et al., 2003). Химический анализ показал, что мутант Arabidopsis hth имеет уровни парафина дикого типа, но ненормальное количество и состав кутина. В частности, он снизил уровни дикарбоновых кислот и увеличил количество ω-гидроксикислот, что привело авторов к предположению, что HTH может играть роль в окислении ω-гидроксижирных кислот до мономеров кутина дикарбоновых кислот, которые характерны для стебля Arabidopsis. кутикулы листьев (Курдюков и др., 2006б). Поскольку мономеры кутина дикарбоновой кислоты необычно многочисленны у Arabidopsis, будет интересно посмотреть, являются ли связанные с HTH белки столь же важными для образования кутикулы у других видов, у которых этот класс мономеров недостаточен.
Другой пример «сиротского» белка, связанного с кутикулой, получен в результате анализа мутанта Arabidopsis bodyguard ( bdg ), который демонстрирует микроскопически дезорганизованную кутикулу с повышенной проницаемостью, но значительно повышенными уровнями воска и кутина (Курдюков и др. ., 2006а). Белок BDG имеет сходство последовательности с белками семейства α / β-гидролаз, но о ферментативной активности не сообщалось. Белок локализован во внешней клеточной стенке эпидермиса ниже кутикулы, что привело авторов к предположению, что BDG может участвовать в полимеризации кутина, хотя повышенное количество полимерного кутина в мутанте может служить аргументом против этого (Курдюков и др., 2006а). Мутация BDG3 , близкого гомолога BDG , приводит к дезорганизации цветочных наногребней, структур эпидермиса лепестков, которые состоят из кутина (Shi et al., 2011). Более того, было показано, что ключевые факторы транскрипции кутина SHN1, SHN2 и SHN3 активируют промотор BDG3 (Shi et al., 2011). Взятые вместе, эти результаты убедительно указывают на то, что белки BDG тесно связаны с образованием полимера кутина, хотя механизм их действия остается загадочным.
Наконец, дефект в формировании цветочных наногребней был также идентифицирован у мутанта Arabidopsis с дефектом кутикулярных гребней ( dcr ), который показал значительный дефицит цветочного кутина, но менее радикальные изменения кутина листа и стебля ( Паникашвили и др., 2009). DCR кодирует белок семейства ацилтрансфераз BAHD, который локализуется в цитоплазме, и было высказано предположение, что он может участвовать в ацильном переносе мономеров кутина с образованием промежуточных соединений-предшественников или олигомерных структур (Panikashvili et al., 2009). Однако позже было проведено биохимическое исследование DCR, и было показано, что он обладает in vitro диацилглицерин-ацилтрансферазной активностью, приводящей к образованию триацилглицерина (Rani et al., 2010). Роль цитоплазматических промежуточных продуктов триацилглицерина в биосинтезе кутина не согласуется с какими-либо известными этапами этого пути, но DCR явно необходим для биосинтеза кутина в органах цветков Arabidopsis. Дальнейшая работа будет необходима для определения нативного субстрата и продукта DCR, чтобы выяснить его роль в биосинтезе кутина.
ФУНКЦИИ КУТИКУЛЫ
Кутикула растения чаще всего связана с обеспечением фиксированного барьера для чрезмерной транспирационной потери воды, позволяя устьицам динамически контролировать газообмен и транспирацию. Однако он развил ряд второстепенных функций, которые согласуются с его местом в качестве самого внешнего слоя первичных надземных органов: он формирует физический барьер, который является первой линией защиты от вредителей и патогенов; у многих видов сложные эпикутикулярные кристаллы помогают формировать самоочищающуюся поверхность, предотвращая попадание пыли и другого мусора на солнечный свет; в некоторых случаях он может экранировать чрезмерный ультрафиолетовый свет; наконец, как определяющий признак эпидермиса, он играет центральную роль в развитии, физически устанавливая границы органов.
Структура кутикулы и водонепроницаемые свойства
Распространено мнение, что толстая кутикула связана с более низкой водопроницаемостью и, следовательно, повышенной устойчивостью к водному стрессу. Однако сравнительные исследования водопроницаемости кутикулы разных видов показали, что нет корреляции ни с толщиной кутикулы, ни с количеством воска (Riederer and Schreiber, 2001). Точно так же количество кутина не обязательно является показателем водной проницаемости кутикулы (CWP).Например, исследования трех мутантов томатов ( cd1 — cd3 ), каждый из которых имеет более чем 95% снижение уровней кутина в плодах, выявили лишь незначительное увеличение скорости потери воды, и даже среди мутантов, находящихся там. не было четкой корреляции между количеством кутина и восприимчивостью к высыханию (Isaacson et al., 2009). Однако дефицит кутина, который ведет к организационным дефектам, может быть вредным для проницаемости кутикулы (Bessire et al., 2011). В отличие от отсутствия связи с кутином, обширное удаление воска из плодов томата, достигнутое путем кратковременного погружения плода в органический растворитель, указывает на то, что воски вносят примерно 95% кутикулярного сопротивления диффузии воды, по крайней мере, в плоды томата (Leide et al., 2007).
Определенные классы соединений, по-видимому, связаны с водонепроницаемыми свойствами кутикулы; в частности, более неполярные компоненты, такие как алканы, имеют тенденцию быть связаны с пониженным CWP, в то время как неалифатические соединения парафина, такие как тритерпеноиды, вероятно, являются менее эффективным водным барьером (Leide et al., 2007; Buschhaus and Jetter, 2012). Это согласуется с моделью, в которой кутикулярные воски локализуются либо в кристаллических, либо в аморфных областях кутикулы, а алифатические соединения образуют кристаллические «плотины», непроницаемые для воды, заставляя воду и другие полярные метаболиты диффундировать обходным путем через аморфные домены, которые образованы более полярными и циклическими парафинами (Riederer and Schreiber, 1995).Идея о том, что пропорция алканов, а не общее количество парафина оказывает наиболее значительное влияние на CWP, была проиллюстрирована недавним исследованием, проведенным на основе обратного скрещивания популяции Capsicum annum и Capsicum chinense , двух видов перца с высоким и низким уровнем воды после сбора урожая. нормы потерь соответственно. В 20 семьях, подвергнутых обратному скрещиванию, CWP обратно коррелировал с количеством алканов в парафине, но не с общим количеством парафина, и более быстро высыхающий родительский элемент имел в три раза больше парафинового покрытия, чем родительский, который демонстрировал низкую потерю воды после сбора урожая (Parsons et al. ., 2012). Таким образом, устойчивость к потере воды в первую очередь приписывается парафину, а не кутину, но нет прямой корреляции между количеством любого из компонентов и CWP. Скорее, похоже, что CWP в первую очередь определяется конкретной смесью внутрикутикулярных и эпикутикулярных восков, а также их упаковкой и организацией в структуре кутикулы.
Эффект лотоса
Поразительной особенностью листьев многих растений является то, что вода имеет тенденцию скатываться в капли и скатываться по земле, собирая и смывая частицы и мусор с поверхности листьев.Эффективность этого механизма самоочищения, называемого «эффектом лотоса», варьируется у разных видов и во время онтогенеза органа, но это коррелирует с обилием эпикутикулярных кристаллов воска, которые отталкивают воду и позволяют воздуху образовываться под каплями. (Бартлотт и Нейнхейс, 1997). Считается, что эта самоочищающаяся поверхность помогает предотвратить накопление пыли, которая блокирует солнечный свет и замедляет фотосинтез, и что это также может сыграть важную роль в смывании спор патогенов до их прорастания.Несмотря на очевидные преимущества самоочищающейся поверхности, нет четкого примера того, как эта черта дает адаптивное преимущество. С точки зрения фотосинтеза, вероятно, существует компромисс между самоочищающейся поверхностью и повышенной дисперсией света эпикутикулярными кристаллами воска, как обсуждается ниже. Тем не менее, благодаря открытию этого эффекта, поверхности с высокой степенью гидрофобности и микроскопической текстурой были использованы в качестве эффективных биомиметических технических материалов (Bhushan, 2012), а улучшенные самоочищающиеся поверхности сельскохозяйственных культур могут стать продуктивным направлением исследований.
Кутикула как барьер против вредителей и патогенов
Кутикула растения представляет собой физический барьер для патогенов, которые иначе не проникают в растение через устьица, раны или переносчики. Однако было показано, что грибковые патогены разрушают кутикулу, используя комбинацию ферментативной деградации и механического разрыва. Последнее часто достигается за счет образования набухшей структуры аппрессора, которая расширяет инфекционный штифт за счет тургорного давления (Deising et al., 2000). Хотя механического разрыва может быть достаточно для проникновения в кутикулу, особенно в более тонкую кутикулу (Tenberge, 2007), большинство грибковых патогенов также секретируют кутиназы, класс небольших неспецифических эстераз, которые гидролизуют полиэфир кутина и высвобождают свободные мономеры кутина (Longhi and Cambillau, 1999). ). Мономеры кутина, которые высвобождаются во время гидролиза полимерного кутина, могут действовать как элиситоры защитных реакций растений и, таким образом, классифицируются как молекулярные структуры, связанные с повреждениями. В микромолярных концентрациях эти соединения вызывают выработку перекиси водорода и другие защитные реакции (Schweizer et al., 1996; Каусс и др., 1999). Однако механизм восприятия растениями свободных мономеров кутина в настоящее время неизвестен (Boller and Felix, 2009).
Кутин, по-видимому, более важен, чем воск для формирования барьера для проникновения патогенов, хотя не существует последовательной корреляции между количеством кутина и устойчивостью к патогенам. В плодах томата резко сниженные уровни кутина у трех мутантов cd были связаны с повышенной восприимчивостью к инфекции, вызванной поверхностной инокуляцией Botrytis cinerea , а также к условно-патогенным микробам (Isaacson et al., 2009). Однако у Arabidopsis ряд мутантов с дефицитом кутина и растений, эктопически сверхэкспрессирующих кутиназы грибов, проявляют повышенную устойчивость к B. cinerea (Bessire et al., 2007, 2011; Chassot et al., 2007; Tang et al., 2007). В этом случае повышенная проницаемость кутикулы, по-видимому, усиливает диффузию элиситоров, полученных из посевного материала, которые индуцируют продукцию небольших полярных противогрибковых соединений, которые, в свою очередь, ингибируют рост B. cinerea (Bessire et al., 2007).Напротив, мутант lacs2 Arabidopsis и сверхэкспрессоры кутиназы не проявили изменений в их восприимчивости к ряду других грибковых патогенов (Bessire et al., 2007), а мутация lacs2 также увеличила восприимчивость к обычно авирулентному штамму Pseudomonas. syringae (Tang et al., 2007). Таким образом, кутин играет важную роль в качестве физического барьера для многих патогенов, однако крайний дефицит Arabidopsis может привести к повышенной устойчивости к некоторым патогенам посредством вторичного, но не до конца изученного механизма, который включает индукцию защиты растений.Дополнительный уровень сложности был предложен наблюдением, что кутин может индуцировать экспрессию генов у растительных патогенов и, как было показано, индуцирует экспрессию аппрессориума в Colletotrichum trifolii (Dickman et al., 2003). Это подчеркивает конкурирующее давление отбора, направленное на создание и нарушение барьеров для кутикулы на границе поверхности растения (Chassot and Metraux, 2005).
Несмотря на важность кутина во взаимодействиях между растениями и патогенами, первая поверхность, с которой сталкиваются патогены для листвы, образована эпикутикулярными кристаллами и пленками воска.Помимо эффекта лотоса, который способствует смыванию спор с поверхности растения перед прорастанием, есть несколько указаний на то, что эпикутикулярные восковые структуры и состав играют важную роль в определении развития грибковых патогенов и, следовательно, патогенности. Альдегид C26 n- гексакозанил, компонент кутикулярного воска у многих видов Poaceae, может вызывать in vitro образование аппрессория мучнистой росой Blumeria graminis (Tsuba et al., 2002; Ringelmann et al., 2009; Hansjakob et al., 2010). Это наблюдение дополнительно подтверждается исследованиями мутанта кукурузы glossy1 , который не накапливает альдегиды в своем восковом комплементе. B. graminis образование аппрессория существенно снижается на поверхности листа мутанта glossy1 , но может быть восстановлено до нормального уровня путем применения n- гексакозанила (Hansjakob et al., 2011). Другой пример влияния восков на патогенность — это ингибитор прорастания трубок ржавчины1 ( irg1 ) мутант M.truncatula , который демонстрирует уменьшенное количество эпикутикулярных кристаллов воска на абаксиальной поверхности листа, что соответствует значительному уменьшению парафиновых первичных спиртовых групп. Было показано, что это поверхностное изменение снижает дифференциацию спор возбудителей ржавчинных грибов Phakopsora pachyrhizi и Puccinia emaculata и антракноза C. trifolii, , что приводит к невосприимчивости (Uppalapati et al., 2012). Было обнаружено, что ген IRG1 кодирует фактор транскрипции цинкового пальца C 2 H 2 , который ранее был идентифицирован как регулятор морфологии рассеченного листа (Chen et al., 2010). Пониженные уровни транскриптов предполагаемых ортологов MYB96 и CER4 также наблюдались у мутанта irg1 , что согласуется с восковым фенотипом. Значимость восков и кутина в устойчивости к патогенам, таким образом, предполагается в общем смысле, но, как и в случае проницаемости кутикулы, мало что известно об относительной важности конкретных молекулярных классов или их межмолекулярных ассоциаций и упаковки в структуре кутикулы.
Эпикутикулярные воски также могут играть важную роль во взаимодействиях растений и насекомых; действительно, кристаллы эпикутикулярного воска могут образовывать нестабильную поверхность, которая препятствует прикреплению насекомых или их передвижению по поверхности растений (Borodich et al., 2010). Ярким примером этого являются плотоядные растения-кувшины ( Nepenthes spp.), Которые ловят насекомых через скользкую внутреннюю поверхность, покрытую эпикутикулярными кристаллами воска (Riedel et al., 2007). Для более подробного обзора химической экологии кутикулы см. Müller and Riederer (2005).
Кутикула и развитие
Помимо обеспечения физических барьеров для воды и микробов, кутикула, по-видимому, играет важную роль в определении границ органов во время развития, поскольку растения с кутикулой, демонстрирующей повышенную проницаемость и структурные дефекты, часто демонстрируют многочисленные эктопические слияния органов. .Это явление наблюдалось у мутантов томатов с дефицитом воска (Смирнова и др., 2013), ряда мутантов Arabidopsis с аномальной кутикулой (Ефремов и др., 1999; Wellesen et al., 2001; Курдюков и др., 2006a). ; Bird et al., 2007) и трансгенных растений Arabidopsis, сверхэкспрессирующих секретируемую кутиназу грибов (Sieber et al., 2000). Зоны слияния часто отмечены двумя соседними стенками полисахаридных клеток без видимой кутикулы, разделяющей два органа, хотя слитые эпидермальные слои сохраняют свою идентичность, на что указывает дифференциация внутренних нефункциональных устьиц в зонах слияния (Sieber et al., 2000). В каждом из трех мутантов Arabidopsis, демонстрирующих слияние органов, lacerata , bodyguard и fiddlehead , эктопические слияния органов и дефекты кутикулярной проницаемости могут быть частично подавлены второй мутацией в SERRATE (Voisin et al., 2009) . SERRATE представляет собой белок цинкового пальца C 2 H 2 , который необходим для биогенеза микроРНК, а гипоморфные аллели обнаруживают многочисленные дефекты развития, включая зубчатые края листьев (Dong et al., 2008). Хотя механизм действия SERRATE как супрессора слияния кутикулы остается неясным, этот результат предполагает существование пути целостности кутикулы, который интегрирован с программами развития эпидермиса. Идентификация дополнительных супрессоров онтогенетических фенотипов, связанных с мутантами кутикулы, должна быть информативной для выяснения пути целостности кутикулы.
Защита от УФ-излучения
УФ-свет в УФ-В спектре составляет значительную часть дневного света, который достигает земной поверхности, и он может угрожать жизни растений, повреждая ДНК, фотосинтетический аппарат и мембранные липиды (Розема и др. ., 1997). В результате растения разработали ряд стратегий для защиты от УФ-В излучения. К ним относятся разнообразные растворимые флавоноидные пигменты, которые обычно локализуются в вакуолях эпидермальных клеток, фенольные соединения, присутствующие в полисахаридной клеточной стенке, и липофильные фенольные молекулы, ковалентно связанные с кутином или связанные с восками (Pfündel et al., 2006) . Обследование изолированных кутикул у ряда видов животных показало, что в целом эффективное экранирование спектра УФ-В, но неизменно высокий коэффициент пропускания в более высоких длинах волн, которые являются фотосинтетически активными (Krauss et al., 1997). В дополнение к поглощению света кутикула растения может в некоторой степени отражать свет, предположительно, в зависимости от количества кристаллов эпикутикулярного воска. Например, Dudleya brittonnii может отражать до 83% УФ-В, но это значение существенно снижается при удалении эпикутикулярных восков (Mulroy, 1979). Гладкие глянцевые «голые» кутикулы обычно отражают лишь небольшое количество света (менее 10%), но сизые поверхности растений обладают умеренной отражающей способностью и обычно показывают коэффициент отражения примерно от 20% до 30% в УФ и видимом спектрах (Pfündel et al., 2006). Воски отражают как УФ, так и видимый свет, но не обязательно в одинаковой степени, а коэффициент отражения УФ, как сообщается, в некоторых случаях выше (Holmes and Keiller, 2002). Хотя отражение света обеспечивает важный защитный механизм, особенно ограничивая повреждающее УФ-излучение, вероятно, существует компромисс с эффективностью фотосинтеза в условиях, когда интенсивность света ограничена (Pfündel et al., 2006). В этом отношении интересной областью будущих исследований может быть определение того, можно ли прогнозируемо изменить относительные пропорции отражения УФ и видимого света путем изменения состава эпикутикулярных восков.
ЗАКЛЮЧЕНИЕ И ПЕРСПЕКТИВЫ
Как описано выше, некоторые ключевые области биогенеза кутикулы остаются малоизученными. Во-первых, механизм внутриклеточного и внеклеточного транспорта предшественников воска и кутина остается неизвестным, хотя ключевые переносчики ABC, необходимые для их экспорта через плазматическую мембрану, были идентифицированы (Pighin et al., 2004; Bird et al., 2007; Chen et al. ., 2011). Была идентифицирована первая кутин-синтаза (Girard et al., 2012; Yeats et al., 2012b), но, безусловно, существуют дополнительные кутин-синтазы, и еще предстоит выяснить, связаны ли они с CD1 или принадлежат к разным семействам белков.Требуется ли после полимеризации кутина модификация полимерной структуры, чтобы приспособиться к расширению органа? Если да, то какие ферменты участвуют в этом процессе?
Хотя наше понимание биосинтеза кутикулы на молекулярном уровне остается неполным, недавний прогресс в расшифровке этих путей приближает нас, чем когда-либо, к способности выборочно изменять свойства кутикулы для повышения продуктивности сельского хозяйства. Однако способность делать такие модификации рационально потребует понимания сложности функции кутикулы на молекулярном уровне, и в этом отношении был достигнут гораздо меньший прогресс.С этой целью дальнейшая работа, направленная на понимание экофизиологических функций кутикулы в определенных мутантных фонах, а также у генетически поддающихся трактовке диких видов, обеспечит основу для понимания сложного взаимодействия структуры, состава и функции кутикулы (Yeats et al. др., 2012а). Хотя за последнее десятилетие произошел беспрецедентный прогресс в молекулярной биологии биогенеза кутикулы, многие исследования выявили сложности в функции кутикулы, которые подчеркивают тот факт, что кутикула — это гораздо больше, чем просто заранее сформированный барьер для потери воды.
Что такое кутикула волоса?
Чтобы понять, что такое кутикула волоса, вам сначала нужно узнать немного больше об основах анатомии волос (мы обещаем, что это намного интереснее, чем уроки анатомии, которые вы проходили в старшей школе).
Волосы состоят из 3-х основных частей:
кутикулы (защитный слой, о котором мы узнаем сегодня)
кора головного мозга (самый толстый слой, в котором сосредоточена большая часть пигмента). )
продолговатый мозг (очень тонкий внутренний слой — у некоторых волосков его даже нет.)
Волосы устроены таким образом, что кора и продолговатый мозг составляют сердцевину пряди волос. Кутикула окружает и защищает их обоих от внешних факторов стресса.
Эти факторы стресса могут включать воду, солнце, нагревательные инструменты, расчески, щетки, жесткую воду … Есть много вещей, которые могут повредить ваши волосы.
Защита — это большая работа, поэтому кутикула представляет собой сложный слой — он состоит из миллионов и миллионов маленьких чешуек, которые наложены друг на друга. Эти чешуйки направлены вниз, от корней, и имеют форму, ограничивающую трение между отдельными волосками.Вблизи они выглядят как кровельная черепица или чешуя на рыбе и движутся друг против друга, так что ваши волосы могут гнуться, не отслаиваясь.
Кутикула — это самая осязаемая часть ваших волос, и вы можете сами оценить форму чешуек без микроскопа. Ущипните прядь волос и проведите пальцами от корня до кончика. Плавно и легко? Теперь сделайте то же самое, но в противоположном направлении: ущипните кончик волос и потрогайте вверх.Не так весело и как-то пискляво? Когда вы проводите пальцами от кончика к корню, вы натыкаетесь на неровные края чешуйчатых слоев кутикулы — это неприятно.
Толщина кутикулы и расовое наследие
Толстая кутикула обычно означает, что уязвимые части волос (мозговое вещество и кора) лучше противостоят повреждениям. Эта толщина часто имеет отношение к расовому наследию. Например, у людей азиатского происхождения кутикула часто бывает наиболее слоистой, в то время как у людей африканского происхождения кутикула более тонкая. Это одна из причин, почему афротекстурные волосы так подвержены повреждениям, несмотря на то, насколько жесткими кажутся отдельные прядки. — защитный слой не очень толстый.
Как выглядит здоровая кутикула?
Здоровая кутикула — это гладкая закрытая кутикула с полностью неповрежденными чешуйками. Кутикула такого типа дает много блеска, так как ее чешуйки расположены под правильным углом, чтобы отражать наибольшее количество света. Он не спутывается с другими волосами и приятен на ощупь.
Как выглядит поврежденная кутикула?
Поврежденная кутикула грубая и тусклая, с приподнятыми или удаленными чешуйками. Такая кутикула оставляет незащищенной уязвимую сердцевину волосяного волокна. Волосы с поврежденной кутикулой также очень спутаны, поскольку чешуйки, которые обычно направлены вниз, приподняты и вызывают трение.
Можно ли открыть / закрыть кутикулу волоса?
…вид. Раскрыть кутикулу волоса — самая простая задача.Если вы когда-либо окрашивали или обесцвечивали волосы, значит, вы уже успешно открыли кутикулу. Чтобы цвет волос «прилип», он должен проскользнуть под чешуйки в кору.
Вы также немного приоткрываете кутикулу каждый раз, когда моете волосы или даже просто намочите их. Вода и шампунь искажают форму кутикулы, поэтому мы советуем ополаскивать и мыть волосы всего несколько раз в неделю. Но закрытие кутикулы волос? Это сложная часть.Как только кутикула волоса деформируется, не так просто вернуть ее прежнюю форму. Вы можете «приклеить» кутикулу обратно вниз и «залатать» места, где она была удалена, с помощью масел и силиконов в кондиционере, но эти чешуйки, вероятно, не приобретут прежнюю форму сами по себе. Это одна из причин, по которой обесцвеченные / окрашенные волосы требуют большего ухода по сравнению с девственными волосами — эти процессы по своей сути разрушают волосяное волокно и требуют тонны TLC (ограничение чистки, кондиционирование при каждой стирке, ограничение тепловой укладки и т. д.), чтобы противодействовать любым эффективным способом.
Как защитить кутикулу волос?
Для начала вам, вероятно, будет полезно средство для волос, содержащее высококачественный силикон. Силиконы часто неправильно понимают и клевещут, но они являются одними из самых эффективных ингредиентов для ухода за волосами.
Силиконы — это, по сути, броня для ваших волос — они работают, покрывая волосы сверхтонким гидрофобным покрытием. Силикон не только защищает от плохого, но и сохраняет хорошее.Пока, тепловое повреждение кутикулы, привет, задержанная влага.
Вы также можете надевать и снимать их по своему усмотрению; Быстрый душ снимет силиконовый слой с кутикулы STAT.
В наших продуктах мы используем пропокситетраметилпиперидинилдиметикон — длинное название удивительного ингредиента. Это уникальный запатентованный силикон, защищающий волосы от тепла, ультрафиолетовых лучей и других факторов окружающей среды. Он также увеличивает объем, мягкость и блеск волос и действует как поглотитель свободных радикалов — большая победа во всех отношениях.
Мне кажется, у меня повреждена кутикула волос. Что мне делать?
👋Привет! Вы попали в нужное место! Мы начнем с сопоставления вас с вашим личным химиком, а затем начнется путешествие по здоровым волосам. Начните здесь!
Хотите узнать больше о волосах? Вот что стоит проверить дальше:
Полезен ли лимонный сок для волос?
Получите пикантные детали
Изменения текстуры волос 101
Хорошие, плохие и уродливые… когда дело доходит до изменения текстуры волос, мы вам поможем
Может ли тревога вызвать выпадение волос?
Сделайте глубокий вдох … это пойдет на пользу вашим волосам.
Как часто следует чистить волосы?
Мы поговорили со специалистами, чтобы узнать
Жизнь с жирной кожей головы + сухие кончики
# несправедливо, да?
Кутикула — обзор | ScienceDirect Topics
2.7.6 Кутикула и апикальный внеклеточный матрикс
Кутикула является ключевым структурным компонентом трахеи (таблица 8).Кутикула насекомого состоит из фибрилл хитина, β1-4-связанного полимера N -ацетил-d-глюкозамина, встроенного в матрицу белков и липидов (Cohen, 2001), которая обеспечивает серьезный барьер для окружающей среды. Хотя кутикула трахеи непрерывна с кутикулой эпидермиса, ее морфология отличается, поскольку она образует выступающие гребни, называемые тенидиальными складками, которые выступают в просвет (Manning and Krasnow, 1993; Tepass and Hartenstein, 1994). Тенидиальные складки, по-видимому, проходят по спирали вдоль просветной поверхности и, как считается, придают эластичность структуре трахеальных трубок, что позволяет им расширяться и сжиматься во время движений личинки.
Таблица 8. Гены, необходимые для образования кутикулы a
| Ген | Номер CG | Продукт | Экспрессия | Фенотип | Функция | Ссылки |
|---|---|---|---|---|---|---|
| dumpy (dp) | CG15637 | Большой белок внеклеточного матрикса | В трахеальных клетках от стадии ямки до стадии 12 | Структурный дефект кутикулы; трахея не заполняется газом | Предсказанное закрепленное на мембране волокно, поперечно сшитое внутри кутикулы, чтобы обеспечить прочный якорь для подлежащей ткани | Wilkin et al .(2000) |
| зернистая головка (grh) | CG5058 | Фактор транскрипции Elf-1 / NTF-1 | Все клетки трахеи | Тонкая и хрупкая кутикула | Активирует допадекарбоксилат , необходимый для окрашивания и укрепление кутикулы | Bray and Kafatos (1991) |
| krotzkopf verkehrt (kkv) | CG2666 | Хитинсинтаза | Во всех трахеальных клетках, начиная с стадий 11–12 | Катализирует синтез | хитиновые цепи, основной компонент кутикулы | BDGP |
| Матричная металлопротеиназа 1 (Mmp1) | CG4859 | Активность металлоэндопептидазы | Личиночная трахея | У личинок наблюдаются короткие и иногда сломанные DT и другие ветви; неравномерный диаметр DT | Необходим для роста личиночной трахеи и высвобождения кутикулы, может быть из-за неспособности клеток оторваться от кутикулы | Page-McCaw et al .(2003) |
| галечный (peb) = ретроспективный взгляд | CG12212 | Фактор транскрипции цинкового пальца | Все клетки трахеи | Taenidia не образуются на просветной поверхности клеток трахеи | Регулирует сборку taenidia, быть косвенным из-за требований целостности эпителия трахеи | Wilk et al . (2000) |
| Pollux (plx) | CG1093 | Белок внеклеточной адгезии | В первичных ветвях от стадии 14 | Застойная жидкость в трахее у личинок 1-го возраста | Требуется для контакта между эпителием трахеи и внеклеточным матриксом |
BDGP, Berkeley Проект генома дрозофилы ; DT, спинной хобот.
Кутикула трахеи состоит из морфологически различных слоев. Самая апикальная эпикутикула состоит из двух электронно-плотных листочков и покрывает слой прокутикулы, который состоит из хитина и связанных с ним белков. Структура и толщина кутикулярного слоя различны в разных ветвях.
Кутикулярная выстилка трахеи секретируется у позднего эмбриона, начиная со стадии 15 или 16 (Campos-Ortega and Hartenstein, 1985; Tepass and Hartenstein, 1994). На этом этапе сформировалась непрерывная сеть трахеи, но просвет трахеи все еще заполнен жидким и электронно-плотным нитевидным материалом (рис. 7).Кутикула выглядит как неравномерно расположенные электронно-плотные области вдоль апикальной мембраны, связанные с зернистым материалом, откладывающимся в просвете. В течение 1-2 часов после начала секреции формируется кутикулярный бислой (предположительно соответствующий внутренней и внешней эпикутикуле) (Рисунок 7). Тенидиальные складки становятся видимыми, и кутикула созревает к концу стадии 17 (рис. 7).
Рис. 7. Изменения формы трубки и внеклеточных компонентов во время развития дорсального туловища. Чертежи продольных срезов спинного туловища на этапе 14 (слева) и этапе 16 (справа).Дорсальный ствол расширяется примерно в три раза в диаметре от стадии 14 до стадии 16 и активно секретирует апикальные компоненты просвета в течение этого периода времени. На стадии 14 апикальный внеклеточный матрикс (EM) покрывает апикальную клеточную поверхность, и нитчатый материал начинает накапливаться в просвете. На стадии 16 кутикула с характерными тенидиями выстилает апикальную клеточную поверхность, а просвет заполняется фибриллярным материалом. Это очищается на стадии 17 до вылупления личинки.Базальная пластинка связана с внешней поверхностью клетки на протяжении всего трахеогенеза.
Фактор транскрипции домена цинкового пальца, кодируемый геном pebbled ( peb , также называемый hindsight ) (Wilk et al ., 2000), и внеклеточным белком адгезии клеток, Pollux (Plx) (Zhang et al., ., 1996) (Таблица 8) необходимы для сборки taenidia и организации и прикрепления апикального внеклеточного матрикса к эпителию.Регулярное формирование паттерна taenidia генерируется неизвестным механизмом, который может включать регулярное расположение апикальных микровиллярных выступов нижележащего эпителия трахеи.
Синтез хитиновых цепей, катализируемый хитинсинтазами, по-видимому, происходит внутриклеточно. Анализ хитинсинтазы мясной мухи овцы ( Lucilia cuprina ) предполагает наличие интегрального мембранного белка с каталитическим доменом, расположенным на цитозольной стороне плазматической мембраны (Tellam et al., ., 2000). Таким образом, полимеры должны быть экструдированы через мембрану, возможно, через поры, образованные 15-20 предсказанными трансмембранными доменами хитинсинтазы. Были идентифицированы два гена Drosophila , кодирующие хитинсинтазы (Gagou et al ., 2002), и по крайней мере один из них, krotzkopf verkehrt ( kkv ) (Ostrowski et al ., 2002) (Таблица 8 ), экспрессируется в развивающейся эмбриональной трахее. Неожиданно kkv начинает экспрессироваться во всех трахеальных клетках уже на стадии 13, намного раньше, чем сообщалось о секреции кутикулы.Возможно, Kkv продуцируется как зимоген, который сначала активируется на стадии 15; напротив, секреция хитина начинается задолго до появления кутикулярного слоя в препаратах для просвечивающей электронной микроскопии (ПЭМ).
Фактор транскрипции Grainy Head (Grh) (Таблица 8), который экспрессируется во всех трахеальных клетках на протяжении эмбриогенеза, также, по-видимому, участвует в образовании кутикулы, поскольку одним из его генов-мишеней является допадекарбоксилаза ( ddc ) ( Таблица 8), кодирующая фермент, необходимый для затвердевания и потемнения кутикулы (Bray and Kafatos, 1991).Ddc экспрессируется на поздних стадиях эмбриогенеза (стадия 17) и, таким образом, участвует в терминальной стадии программы синтеза кутикулы.
Помимо апикальной кутикулы, клетки трахеи, а также клетки эпидермиса секретируют внутрипросветный матрикс, что очевидно из исследований электронной микроскопии (Tepass and Hartenstein, 1994). Это появляется уже на стадии 11 и может быть замечено до поздней стадии 15, когда кутикула становится очевидной. Молекулярный состав или функция этой матрицы неизвестны. Он может содержать хитин и молекулы, распознаваемые орфанными антителами (например, антиген 2A12), которые окрашивают просвет трахеи, и большой белок Dumpy, который может образовывать большие внутрипросветные фибриллы (Wilkin et al ., 2000). Такой матрикс может обеспечивать якорение для белков, которые не только способствуют перекрестному связыванию и структурной поддержке матрикса, но также влияют на поведение подлежащего трахеального эпителия.
Структура волос
Структура волосЭтот веб-сайт требует файлов cookie для обеспечения всех его функций. Для получения дополнительной информации о том, какие данные содержатся в файлах cookie, пожалуйста, посетите нашу страницу ПОЛИТИКА COOKIES. Чтобы принимать файлы cookie с этого сайта, нажмите кнопку ниже.
Принять и продолжить
Похоже, в вашем браузере отключен JavaScript. Для максимального удобства работы на нашем сайте обязательно включите Javascript в своем браузере.
Анабель Кингсли
Президент бренда
Опубликовано в январе 2016 г.
Каждый из ваших волос вырастает из отдельного волосяного фолликула. Волосяная луковица расположена у основания каждого волосяного фолликула и содержит растущие волосковые клетки.Они постоянно делятся и толкаются вверх, постепенно затвердевая. Достигнув верхней части луковицы, они образуют шесть концентрических слоев. Три внутренних слоя становятся волосами, состоящими из кутикулы, коры и продолговатого мозга (хотя мозговое вещество присутствует не всегда, особенно в волосах с меньшим диаметром). Три внешних слоя становятся выстилкой фолликула.
Определенные клетки волосяной луковицы, называемые меланоцитами, производят пигмент меланин, который придает цвет волосам.
Форма волосяного фолликула определяет форму ваших волос (т. Е. Прямые, волнистые или вьющиеся).
Стержень волоса — это часть волос, которая видна над кожей головы. Он состоит из белка, называемого кератином, уплотненного и слитого вместе.
Кератин — это тот же материал, из которого состоят ногти, копыта, перья и когти.
Кератин — чрезвычайно сильный белок, очень устойчивый к износу. Он удерживается вместе двумя типами связей: дисульфидными связями и водородными связями.
Дисульфидные связки чрезвычайно прочные. Фактически, они являются одними из самых сильных естественных связей в мире. Когда вы делаете химическую завивку или расслабляете волосы, эти дисульфидные связи разрываются и возвращаются к другой конфигурации. Это то, что позволяет навсегда изменить форму волос.
Водородные связи слабее и многочисленнее, чем дисульфидные связи. Они помогают придать волосам гибкость. Когда вы намочите волосы, они легко ломаются, и их можно временно восстановить с помощью тепла, пока они снова не станут влажными (из-за воды или влажности).Это то, что позволяет укладывать волосы феном и щипцами после мытья.
Стержень вашего волоса состоит из трех слоев:
Кутикула
Кутикула — это защитный слой ваших волос, состоящий из перекрывающихся ячеек, таких как рыбья чешуя или черепица, но обращенных вниз. Здоровая кутикула гладкая и плоская. Это придает блеск волосам и защищает внутренние слои от повреждений. Он также сводит к минимуму перемещение влаги внутрь и наружу коры головного мозга, таким образом поддерживая баланс гидратации и гибкость ваших волос.Однако химические процессы и атмосферные воздействия могут приподнять кутикулу и нарушить этот баланс, в результате чего волосы станут сухими и ломкими.
Cortex
Кора головного мозга формирует основную массу и пигмент (цвет) ваших волос. Он состоит из длинных кератиновых волокон, которые скрепляются дисульфидными и водородными связями. Здоровье коры головного мозга во многом зависит от целостности защищающей ее кутикулы.
Медулла
Мозговое вещество присутствует только в более густых типах волос и является самым внутренним слоем ваших волос.Он состоит из мягкой, тонкой сердцевины из прозрачных ячеек и воздушных пространств.
Вас также может заинтересовать:
Купите наши средства для ухода за структурой волос для сильных прядей
Мы не можем найти продукты, соответствующие вашему выбору.
{{/ thumbnail_url}} {{{_highlightResult.name.value}}}{{#categories_without_path}} в {{{category_without_path}}} {{/ category_without_path}} {{#_highlightResult.цвет}} {{# _highlightResult.color.value}} {{#categories_without_path}} | {{/ category_without_path}} Цвет: {{{_highlightResult.color.value}}} {{/_highlightResult.color.value}} {{/_highlightResult.color}}
{{price.USD.default_formated}} {{# price.USD.default_original_formated}} {{цена.USD.default_original_formated}} {{/price.USD.default_original_formated}} {{# price.USD.default_tier_formated}} {{price.USD.default_tier_formated}} {{/price.USD.default_tier_formated}}
© Philip Kingsley Products Ltd., 2021 г. Все права защищены.
Динамика диффузии и адсорбции влаги в кутикуле растений, включая роль целлюлозы
Модельный каркас
Мы описываем комплексную механистическую модель для изолированного переноса воды кутикулой.Модель представляет собой нелинейную одномерную диффузионную модель, включающую уравнения в частных производных. Мы кратко опишем формулировку моделирования, которая основана на предыдущих работах авторов 3,17,18 , и мы отсылаем читателя к этим работам для полного описания вспомогательных уравнений. (5), (8) и (9) и дополнительные уравнения. (2) и (3).
Водные поры переносят воду, и их свойства различаются у разных видов растений, с относительной влажностью, и меньше подвержены влиянию температуры и присутствия восков, чем липофильный путь 6,27 .Сорбционная способность целлюлозы выше, чем у других материалов для кутикулы, и она неравномерно распределена 30,33,34 . Мы моделируем целлюлозу как отдельный путь от водных пор, что является разумным из-за результатов, показанных на дополнительном рис. 1. Мы учитываем воду, которая может перемещаться тремя путями: водные поры (A), липофильный путь (L), и целлюлоза (C). Эти три пути регулируются разными механизмами, поэтому их необходимо моделировать отдельно. Концентрация влаги изменяется в основном по глубине мембраны кутикулы, z (0 ≤ z ≤ b ).Влага внутри кутикулы считается жидкой водой, что является разумным предположением, основанным на экспериментальных данных 13,28 . Эта модель станет дополнением к простой модели диффузии, приспособив уникальные пути кутикулы. Мы включаем важные управляющие механизмы набухания водных пор, климатические условия, такие как температура и относительная влажность, которые влияют на набухание и адсорбцию пор, параметры, которые учитывают различия в видах растений, включая пористость и извилистость, плотность и радиус пор, толщину кутикулы и связывание с поверхностью кутикулы за счет целлюлозы.Модель, как описано в уравнениях. (1) — (13), Дополнительные уравнения. (2) — (8), параметры в таблице 1 и константы в дополнительной таблице 3, для диффузии и адсорбции воды тремя путями кутикулы, следующие:
$$ \ left ({\ varepsilon} _ {{{{\ rm C}}}} + {\ varepsilon} _ {{{{{\ rm L}}}} \ right) \, \ frac {\ partial c} {\ partial t} + \ frac {\ partial \ left ({\ varepsilon} _ {{{{\ rm D}}}} c \ right)} {\ partial t} = \ left ({\ varepsilon} _ {{{{\ rm C}}}}} \, {D} _ {{{{\ rm C}}}} + {\ varepsilon} _ {{{{\ rm L}}}} \, {D} _ {{{{\ rm L}}} } \ right) \ \ frac {{\ partial} ^ {2} c} {\ partial {z} ^ {2}} + \ frac {\ partial} {\ partial z} \ left ({D} _ {{ {{\ rm A}}}} \ frac {\ partial ({\ varepsilon} _ {{{{\ rm D}}}} c)} {\ partial z} \ right) \\ + S, \, \ , 0 \, <\, z \, <\, b, \, t \,> \, 0, $$
(1)
$$ S = — \ frac {2} {{r} _ {{{{\ rm A}}}}} \ left (1 — {\ varepsilon} _ {{{{\ rm D}}}} \ right) \ frac {\ partial {{{\ Gamma}}} _ {{{{{\ rm A}}}}} {\ partial t} — {\ rho} _ {{{{{\ rm C}}} } \, \ left (1 — {\ varepsilon} _ {{{{\ rm C}}}} \ right) \ frac {\ partial {{{\ Gamma}}} _ {{{{\ rm C}} }}} {\ partial t}, $$
(2)
, где c — концентрация жидкой воды в кутикуле, ε D , ε C и ε L — пористость, t — время, D A , D C и D L — коэффициенты диффузии, z — глубина кутикулы, r A — радиус водной поры, Γ A, C — концентрация адсорбированной воды в водных порах и целлюлозе, S — член адсорбции, а ρ C — плотность целлюлозных волокон.Индексы A, D, C и L означают водные поры (A — вся водная пора, а D — водная пора, доступная для диффузии), целлюлозу и липофильный путь. Параметры, которые изменяются в пространстве и времени, включают c ( z , t ), D A ( z , t ), r A ( z , t ), ε D ( z , t ), Γ A, C ( z , t ), как описано в уравнениях.(5), (8), (9), (10) и (11).
Таблица 1 Параметры модели.Экспериментальная установка 4 , которая используется для разработки модели, включая домен (кутикулу) и начальные условия, состоит из астоматозных изолированных кутикулы плодов томата, помещенных в ряд RH. Кутикула изначально сухая, и относительная влажность постепенно увеличивается. Кутикулы взвешивают, и увеличение веса по сравнению с сухой массой регистрируют в двух разных экспериментах, один в почти установившемся состоянии через 6 часов в большом диапазоне влажности, а другой — в течение 10 минут и небольшом выборе влажности. .Оба эксперимента проводятся при постоянной температуре 20 ° C. Влага проникает в кутикулу как с внешней, так и с внутренней поверхности кутикулы. На границах (поверхностях кутикулы) вода проходит в кутикулу и адсорбируется на поверхностях кутикулы из-за присутствия целлюлозы, где вода задерживается и больше не доступна для диффузии, но может увеличить вес кутикулы. Затем вода попадает в кутикулу и может пройти по трем путям. В водных порах свободные молекулы воды могут проходить через кутикулу посредством пассивной диффузии Фика или адсорбироваться монослоем на стенках водных пор, вызывая их набухание.В целлюлозе свободная вода проходит через кутикулу посредством пассивной диффузии Фика или адсорбируется внутри кутикулы на целлюлозных волокнах и на поверхности кутикулы. В липофильном пути вода может диффундировать через кутикулу и находится под влиянием изменения температуры. В формуле. (1), последний член описывается в формуле. (2) адсорбции в водных порах кутикулы и целлюлозе. Термин, регулирующий адсорбцию на водных порах в монослое, сформулирован путем включения площади поверхности поры по объему пор, включая пористость, а в рецептуре целлюлозы вместо этого используется плотность целлюлозы с многослойной адсорбцией.Как свободная, так и адсорбированная вода увеличивают вес кутикулы и требуются для расчета Δ w , увеличения веса кутикулы по сравнению с сухой массой, в дополнительном уравнении. (7). Изотермы адсорбции для Γ A и Γ C описаны более подробно в формулах. (9) и (10).
Граничные условия для механизмов поверхности кутикулы, участвующих в сорбции, имеют большое значение. Экспериментальная установка для кутикулы такова, что RH применяется как на внешних, так и на внутренних граничных условиях.Внешняя и внутренняя поверхности кутикулы в точках z = 0 и z = b регулируются следующими граничными условиями:
$$ — {\ left. \ Left [\ left ({\ varepsilon} _ {{{{\ rm C}}}} {D} _ {{{{\ rm C}}}} + {\ varepsilon} _ {{{{\ rm L}}}}} {D} _ {{{ {\ rm L}}}} \ right) \ frac {\ partial c} {\ partial z} + {D} _ {{{{{\ rm A}}}} \ frac {\ partial ({\ varepsilon} _ {{{{\ rm D}}}} c)} {\ partial z} \ right] \ right |} _ {z = 0} \\ = -h \, \ left ({c} _ {\ infty} -c (0, t) \ left (1- \ frac {c (0, t)} {{c} _ {{{{{{\ rm H}}}}} _ {2} {{{\ rm O}) }}} ^ {{{{{\ rm pure}}} \,}} \ right) \ right) — {k} _ {1} \, c (0, t), \ quad t \; > \; 0, $$
(3)
$$ — {\ left.{{{{\ rm pure}}} \,} \) — концентрация воды как функция температуры, а k 1 и k 2 — константы скорости связывания воды с целлюлозой на поверхности внешняя и внутренняя поверхности кутикулы. Уравнения (3) и (4) являются граничными условиями типа Робена. Уравнения (3) и (4) аналогичны закону нагрева Ньютона и показывают, что поток влаги через границу пропорционален разнице между концентрацией атмосферного пара и уровнями влажности на поверхности кутикулы.Мы включили формулировку логистического роста, которая предотвращает концентрацию воды, превышающую концентрацию чистой воды, что необходимо из-за значений констант в уравнениях. Вода может адсорбироваться на целлюлозе в кутикуле 32,33,34 и последний член уравнений. (3) и (4) описывает адсорбцию целлюлозы на поверхности кутикулы. Эти термины показывают, что адсорбция на поверхности увеличивается пропорционально концентрации влаги на поверхности. Параметры k 1 и k 2 , константы скорости адсорбции воды целлюлозой на поверхностях кутикулы, являются функциями RH и количества присутствующей целлюлозы, как описано в дополнительном уравнении.(5) и дополнительная таблица 3. Внешняя и внутренняя поверхности похожи, за исключением того, что они содержат разное количество целлюлозы, поэтому разные количества воды могут связываться, как описано в дополнительном уравнении. (5) и дополнительная таблица 3.
Проверка модели
Модель, как описано в уравнениях. (1) — (13) и дополнительные уравнения. (2) — (8) решается численно. Затем мы проверяем модельное решение по экспериментальным данным 4 . Мы проверяем модель на двух наборах результатов, один с диапазоном относительной влажности, а второй с выбором влажности и времени.На рисунке 1 показаны результаты модели (зеленые точки) в сравнении с экспериментальными данными в диапазоне относительной влажности. Из-за изменчивой природы кутикулы мы сравнили результаты модели с результатами трех экспериментов на изолированных кутикулах плодов томата с аналогичной экспериментальной установкой (белые кружки) 4,10,12 и аппроксимировали их данные (сплошные кривые) с простое уравнение, как описано в дополнительной таблице 2. На рис. 1 мы видим, что модель хорошо сравнивается со всеми тремя экспериментами в диапазоне влажности.Luque et al. 12 набор данных включен, поскольку он включает высокую влажность, но модель основана на Chamel et al. 4 экспериментальная установка, поэтому по всем значениям влажности модель хорошо сравнивается с Chamel et al. Модель значительно увеличивается (выше линейного тренда) при очень высокой влажности, при относительной влажности 90% и относительной влажности 99%, что является желаемым результатом. Эта тенденция при высокой влажности в значительной степени является результатом прикрепления воды к целлюлозе на внешней и внутренней поверхностях кутикулы, что обсуждается позже (см. Ниже).
На рис. 2 мы видим, что модель хорошо сравнивается с экспериментальными данными при относительной влажности 60% с течением времени, а изменение сухой массы увеличивается пропорционально относительной влажности. Первоначальное быстрое увеличение в значительной степени вызвано целлюлозой и водными порами с меньшим влиянием липофильного пути, обсуждаемого ниже. На рис. 3 мы видим график концентрации молекул воды в выбранные моменты времени с глубиной внутри кутикулы. Первоначально (оранжевый) концентрация воды в кутикуле низкая и постоянная.Затем, по мере увеличения времени, концентрация на границах увеличивается, а затем постепенно диффундирует в кутикулу. В поздние сроки кутикула приближается к концентрации чистой воды. Отметим, что только на рис. 3 показаны диффундирующие молекулы воды, а не адсорбирующиеся молекулы. Из-за асимметричных граничных условий концентрация воды на двух границах одинакова, но не идентична.
Рис. 2: График проверки по сравнению с экспериментальными данными (данные Exp выделены черным цветом) при относительной влажности 60%.Можно увидеть раствор модели кутикулы с относительной влажностью 60% (зеленовато-коричневый), точно соответствующий эксперименту, выделенный черным цветом.(v) представляет решение, используемое для проверки. Общее время эксперимента здесь составляет 10 минут. Мы воспроизводим экспериментальные данные, как описано в Дополнительном уравнении. (1).
Рис. 3: Результаты моделирования кутикулы растения при одной относительной влажности 60% RH для концентрации диффундирующих молекул воды в течение 6 часов.Результаты отображаются в разное время по всей глубине кутикулы. Начальное условие показано оранжевым цветом, а граничные условия для внешней и внутренней поверхностей кутикулы расположены на глубине кутикулы 0 и 1.87 × 10 −5 м соответственно.
Мы проводим анализ чувствительности модели на основе результата валидации (v) на рис. 2 при относительной влажности 60%, сохраняя все параметры одинаковыми, а затем применяя однофакторный метод для определения чувствительности параметров модели, как показано на рис. 4. Выбор чувствительности обсуждается ниже. На рисунках 1 и 2 уже показана чувствительность RH и подтверждена экспериментальными данными. На рис. 4a мы видим результаты изменения F s , фрактальной масштабной размерности, аналогичной извилистости трех путей.Увеличение веса кутикулы очень чувствительно к извилистости проводящих путей, так как это изменяет диффузию. Это единственная полученная обратная зависимость: увеличение F s означает более извилистый и длинный путь, который труднее пересечь, замедляя и ограничивая диффузию и диффузию трех путей. При применении этого результата к экспериментам с изолированной кутикулой извилистость водных пор будет значительно различаться между кутикулами растений из-за анатомии растений, такой как пластинчатая структура, толщина, ориентация пор и возраст растения 5,21,41 .Следовательно, значительный эффект F s указывает на то, что изменение видов растений само по себе оказывает значительное влияние на водный транспорт, и это видно экспериментально 5 .
Рис. 4: Анализ чувствительности модели по сравнению с результатами проверки (v) и экспериментальными данными (данные Exp выделены черным цветом) в качестве контрольной точки, все выполнено при относительной влажности 60%. {{{{{\ rm max}}} \,} \), максимальный радиус водных пор.Обратите внимание на то, что для всех рисунков зависимость прямо пропорциональна, за исключением F s , которая имеет обратную зависимость.На рис. 4b показана чувствительность ε C , пористость целлюлозы. Этот параметр имеет сильную прямую связь с увеличением массы и показывает аналогичные (но прямые, а не обратные) тенденции к F s , поскольку эти параметры изменяют эффективный коэффициент диффузии целлюлозы (уравнение (6)). Чем больше значение пористости, тем больше пор для диффузии, поэтому эта тенденция соответствует ожиданиям.{{{{\ rm max}}} \,} \), максимальный радиус водных пор. Существует прямая взаимосвязь, и увеличение максимального размера пор способствует более быстрой диффузии и адсорбции в порах. Максимальный радиус водной поры имеет несколько существенное влияние на изменение увеличения массы кутикулы. Значение этого эффекта изменения этого параметра для водных пор состоит в том, что водные поры действительно вносят некоторый вклад в общее увеличение массы, но значительный вклад также вносит пористость целлюлозы, наблюдаемая при сравнении изменения массы между максимальным и минимальным значениями на рис. .4б, д.
Дополнительные изученные параметры включают коэффициент скорости связывания целлюлозы на границах, k , пористость липофильного пути, ε L и температуру, T . На дополнительных фиг. 2 и 3, константа скорости связывания при граничных условиях из-за целлюлозы, k , очень мало влияет на увеличение массы в краткосрочной перспективе, но оказывает большое влияние в течение длительного времени. Это указывает на то, что связывание происходит в течение длительного периода времени и слишком медленно по сравнению с масштабом времени диффузии, и что связывание целлюлозы в течение длительного периода времени вносит значительный вклад в результаты, показанные на рис.1. Здесь очевидны две шкалы времени: короткая шкала времени диффузии и адсорбции, определяемая в основном диффузией и адсорбцией целлюлозы и водных пор внутри кутикулы, и более длительная шкала времени, определяемая адсорбцией на целлюлозе на границах, и эти две шкалы времени отражаются обеими эксперименты и математические модели 3,4,18 . Механизм связывания на поверхности кутикулы целлюлозой имеет решающее значение для включения в модель сорбции воды, чтобы мы могли подтвердить результаты на рис.1, и должен быть включен, как и в предыдущих моделях кутикулы 3,17,18 . Важность поверхностного связывания воды целлюлозой также была обнаружена в хорошо известной литературе 35 . На дополнительном рисунке 4 пористость липофильного пути, ε L , оказывает слабое влияние на увеличение массы, что подразумевает, что липофильный путь в некоторой степени способствует увеличению массы, но не в значительной степени в этом исследовании. На дополнительном рисунке 1 экстракция парафина имела лишь небольшой эффект 4 , и результаты, полученные в другом месте 5 , согласуются с тем, что вода перемещается в основном по водному пути и меньше по липофильному пути.Мы отмечаем, что в некоторых исследованиях 10,11 использовались другие методы, химические вещества, предварительные обработки и адъюванты, которые могут повысить роль липофильного пути в проникновении воды. {{{{\ rm max}}}} \) и η A .Параметры, относящиеся к целлюлозе, включая ε C , k и параметры, относящиеся к k и Γ C , также сильно влияют на сорбцию в модели, и это согласуется с экспериментальными данными, так как целлюлоза поглощает значительное количество воды кутикулой 4,31 . RH также имеет большое влияние на увеличение массы. Наши результаты анализа чувствительности возможны. Принимая во внимание все результаты анализа чувствительности, мы обнаруживаем, что целлюлоза и водные поры способствуют увеличению массы кутикулы за более короткий промежуток времени (10 мин), в то время как липофильный путь вносит свой вклад в небольшой степени.В более длительном масштабе времени (6 часов) диффузия в целлюлозе и связывание с целлюлозой на границах приводит к увеличению массы.
Кутикула | Encyclopedia.com
Oxford
просмотров обновлено 29 мая 2018cu · ti · cle / ˈkyoōtikəl / • п. 1. Наружный слой живой ткани, в частности: ∎ Бот. & Zool. защитный восковой или твердый слой, покрывающий эпидермис растения, беспозвоночного или панциря. ∎ внешний клеточный слой волоса. ∎ Зоол. еще один термин для эпидермиса.2. омертвевшая кожа у основания ногтя или ногтя на ноге. ПРОИЗВОДНЫЕ: cu · tic · u · lar / kyoōˈtikyələr / прил.
The Oxford Pocket Dictionary of Current English
oxford
просмотров обновлено 27 июня 2018 г. кутикула1. (в ботанике) Непрерывный восковой слой, покрывающий надземные части растения. Состоящий в основном из кутина, он секретируется эпидермисом, и его основная функция — предотвращать потерю воды.
2. (в зоологии) Слой рогового неклеточного материала, покрывающий и секретируемый эпидермисом многих беспозвоночных. Обычно он состоит из коллагеноподобного белка или хитина, и его основная функция — защита. У членистоногих он также достаточно силен, чтобы действовать как скелет (см. Экзоскелет), а у насекомых он снижает потерю воды. Рост возможен линькой кутикулы (см. Шелушение).
Биологический словарь
Оксфорд
просмотров обновлено 8 мая 2018 г. кутикула1. Непроницаемое покрытие внешних стенок эпидермальных клеток надземных органов растений, состоящее из кутина, кутана или их смеси.
2. Внешний слой насекомого, секретируемый клетками эпидермиса; он имеет сложную структуру, которая варьируется в зависимости от вида. См. СКЕЛЕТНЫЙ МАТЕРИАЛ.
Словарь наук о Земле АЙЛСА АЛЛАБИ и МАЙКЛ АЛЛАБИ
оксфорд
просмотров обновлено июн 08 2018 кутикула (прил. Кутикула) Слой, покрывающий и секретируемый эпидермисом.У беспозвоночных он в основном защищает от механических или (у эндопаразитов, см. ПАРАЗИТИЗМ) химических повреждений. Смотрите EXOSKELETON.Зоологический словарь МАЙКЛ АЛЛАБИ
оксфорд
просмотров обновлено 14 мая 2018 г. кутикула Открытый внешний слой животного. У людей это относится к эпидермису, особенно к омертвевшей коже на краях пальцев. В ботанике это восковой слой на внешней поверхности клеток эпидермиса листьев и стеблей сосудистых растений.Это помогает предотвратить чрезмерную потерю воды.World Encyclopedia
Oxford
просмотров обновлено 17 мая 2018 г. кутикула У растений — тонкий восковой защитный слой, покрывающий поверхность листьев и стеблей. У животных — слой, покрывающий и секретируемый эпидермисом. У беспозвоночных он в основном защищает от механических или (у эндопаразитов, см. Паразитизм) химических повреждений.Экологический словарь МАЙКЛ АЛЛАБИ
оксфорд
просмотров обновлено 17 мая 2018 г. кутикула ( кью -ti-kŭl) n.1. эпидермис кожи.
2. слой твердого или полутвердого материала, который секретируется эпителием и покрывает его.
3. слой клеток, например внешний слой клеток в волосе.
Словарь медсестер
Оксфорд
просмотров обновлено 11 июня 2018 кутикула эпидермиса. XVII. — L. cutīcula , разм. кутис ; см. КОЖАНЫЙ.Краткий Оксфордский словарь английской этимологии T.F. HOAD
Oxford
просмотров обновлено 27 июня 2018 кутикула Тонкий восковой защитный слой, покрывающий поверхность листьев и стеблей растений.Словарь наук о растениях МАЙКЛ АЛЛАБИ
Кутикула волос: понимание этой важной части ваших волос
Кутикула волос. Мы слышали о них раньше, но большинство из нас может даже не знать, что они из себя представляют. Они там, где волосы выходят из головы? Они такие же, как волосяные фолликулы? Проще говоря, кутикула волоса — это самый внешний слой пряди волос.Это то, что защищает волосы от повреждений и придает блеск.
С научной точки зрения Национальный центр биотехнологической информации (NCBI) определяет кутикулу волоса как слой мертвых клеток, которые образуют защитный слой вокруг пряди. Кутикула состоит из чешуйчатых клеток, которые перекрывают друг друга и защищают другие слои, составляющие одну прядь волос. Итак, чтобы ответить на эти предыдущие вопросы: волосяные фолликулы расположены глубоко под кожей головы, а кутикула волоса — это самый внешний слой пряди волос.
Поскольку кутикула — это самый внешний слой волос, их блеск, влажность и гладкость (или их отсутствие) — вот то, что выставлено на всеобщее обозрение. Таким образом, забота о кутикуле волос с помощью правильных продуктов и методов ухода является важной частью поддержания здоровья волос.
Давайте разберем структуру кутикулы наших волос, чтобы узнать, что делает кутикулу здоровой или нездоровой, как это связано с пористостью волос и как улучшить здоровье кутикулы волос.
Какова структура волос?Чтобы лучше продемонстрировать, что такое кутикула волос, нам нужно немного углубиться в науку, чтобы понять структуру отдельной пряди волос. (Не волнуйтесь, вас не будут оценивать по этому поводу.)
Каждое волокно волоса состоит из трех слоев. Самый внутренний слой называется мозговым веществом, который состоит из неплотно упакованных клеток в центре одной нити. Средняя часть волосяного волокна, окружающая мозговое вещество, называется корой и состоит из белков кератина и структурных липидов.Кора составляет большую часть массы прядей волос и окружена слоем кутикулы, который защищает общую структуру каждой пряди.
Хотя одна прядь волос состоит из трех слоев, человеческий глаз видит только кутикулу. Это похоже на защитный экран, который следит за тем, чтобы другие более хрупкие слои оставались неповрежденными. Гладкие и блестящие у вас волосы или вьющиеся и тусклые — это показатель здоровья вашей кутикулы.
Каждая прядь имеет стержень и корень волоса.Стержень волоса — это участок, который заметно выступает из кожи головы. Корень — это часть пряди, которую мы не видим, поскольку она находится внутри кожи головы и простирается до более глубоких слоев кожи.
Волосяной фолликул — это полость в коже черепа, которая удерживает каждый волос на месте. Мышцы, связанные с этими волосяными фолликулами, заставляют каждый волос встать дыбом. Волосяной фолликул состоит из множества нервов, поэтому ваша голова так чувствительна к температуре, трению и волочению за волосы. Когда волосы выдергиваются, из пустого фолликула формируется новый волос, и цикл роста волос восстанавливается.
Как выглядит Кутикула здоровых волос ?Теперь, когда вы знаете немного больше о том, что такое кутикула волоса, вы можете задаться вопросом, как определить, здорова ли кутикула волоса или нет. Здоровая кутикула должна выглядеть гладкой и легко отражать свет. Он также должен выдерживать и ограничивать трение, возникающее при трении стержней волос друг о друга.
Как правило, кутикула здорового волоса будет однородной по текстуре от корня до кончика каждой пряди.Если вы наблюдаете за отдельной прядью и видите, что она такая же гладкая и толстая на конце, как и в начале, это хороший показатель того, что у вас здоровая кутикула. Но если вы заметили, что текстура ваших волос становится тоньше и менее однородной по мере того, как вы доходите до кончика волос, возможно, вы имеете дело с нездоровой кутикулой волос.
Здоровые волосы состоят из тонких слоев клеток, которые накладываются друг на друга, как черепица. Но когда волосы повреждены, опоясывающий лишай раскрывается и становится неровным. Если вы когда-нибудь видели прядь волос под микроскопом, вы поймете эту аналогию.Нездоровые пряди волос похожи на распутанный кусок веревки.
Частые химические обработки и тепловые повреждения от инструментов для укладки являются частыми виновниками повреждения кутикулы. В результате у вас могут возникнуть такие проблемы, как завивка, сухость, ломкость и секущиеся кончики. (Эти типы проблем с волосами более распространены среди людей, которые обычно изменяют свои естественные волосы.) Плохой уход за волосами и воздействие окружающей среды также могут изменить текстуру стержней волос и способствовать ослаблению общей структуры волос.
Но как эти привычки влияют на кутикулу ваших волос? Чтобы химически изменить волосы с помощью таких процессов, как окрашивание, выпрямление или химическая завивка, в коре головного мозга должна произойти химическая реакция. Эти химические вещества должны открыть защитный слой кутикулы, чтобы проникнуть во внутренний слой коры.
Кутикулы волос естественным образом покрыты маслянистым веществом, называемым кожным салом, которое предотвращает высыхание волос. Таким образом, когда корка волос проникает внутрь, защитный кожный жир удаляется.Кутикула открыта и ее необходимо закрыть, чтобы восстановить ее естественное здоровое состояние.
Общие сведения о пористости волосЕще одним показателем здоровья кутикулы является пористость волос, которую Нью-Йоркское общество косметических химиков определяет как степень поглощения или удержания воды вашими волосами, средства для ухода за волосами и средства для ухода за волосами. Если вашим волосам трудно удерживать или впитывать воду и средства для волос, они не смогут должным образом поддерживать кутикулу волос.Есть три типа пористости волос.
Низкая пористостьВолосы с низкой пористостью имеют тенденцию сопротивляться впитыванию воды и химической обработки. Также обычно требуется много времени для высыхания, легко происходит накопление продукта, а не абсорбция, и он менее чувствителен к нагреву или химической обработке.
Типы волос с низкой пористостью: Вьющиеся, кудрявые или химически выпрямленные волосы с грубой структурой.
Наконечники для волос с пористостью : Если у вас пряди с низкой пористостью, вам необходимо восстановить водный баланс волос и обработать волосы надолго.Использование маски для волос с такими ингредиентами, как экстракт саке и эфирное масло жожоба, может помочь увлажнить кутикулу волос.
Средняя ПористостьВолосы средней пористости достаточно хорошо впитывают и удерживают воду. Он легко принимает химические обработки с предсказуемыми результатами.
Типы волос со средней пористостью: Это идеальная пористость волос. Если вам повезло, ваши кутикулы в отличной форме.
Высокая ПористостьВолосы с высокой пористостью — это сильно обработанные волосы, поврежденные химическими веществами или физическим стрессом.Он быстро намокает и сохнет, легко завивается и ломается, а в целом оставляет ощущение сухости. Этот тип пористости очень чувствителен к краскам для волос и имеет тенденцию легко впитывать влагу, но не может ее удерживать.
Типы волос с высокой пористостью: Вьющиеся, сухие или тусклые волосы, которые могут быть повреждены чрезмерным окрашиванием или химической обработкой.
Наконечники для волос с высокой пористостью : Если у вас волосы с высокой пористостью, вам необходимо увлажнить и защитить пряди от завивки и ломкости.Использование легкой сыворотки для волос с такими ингредиентами, как аргановое и подсолнечное масло, может помочь вам добиться гладкой, насыщенной влагой текстуры.
Здоровая кутикула = счастливые волосыЗдоровье кутикулы волос — это поддержание влаги в прядях волос и минимизация повреждений. Это означает, что нужно отказаться от нагрева или химической обработки прядей и обратить внимание на баланс влаги в волосах. Это также означает определение уникальной пористости ваших волос. Если ваши волосы либо чрезмерно, либо недостаточно впитывают продукты (но, кажется, никогда не получают от них никакой пользы), возможно, пришло время начать использовать средства по уходу за волосами с ингредиентами, предназначенными для ваших конкретных целей и потребностей волос.
Например, шампунь и кондиционер Function of Beauty полностью настраиваются в соответствии с вашим типом волос, потребностями и целями. Итак, хотите ли вы питать корни, защитить цвет или придать волосам блеск и блеск, у нас есть формула для вас.
Думайте о кутикуле волос как о хранителе ваших волос. Его защитные свойства защищают внутренние слои каждой пряди от повреждений и придают волосам блеск и гладкость. Хотя окрашивание и химическая обработка волос влияют на здоровье кутикулы, существует множество способов обработать локоны и управлять уникальной пористостью волос, чтобы они оставались в идеальной форме.